Гибридизация ящериц подрода Archaeolacerta и ее эволюционное значение
Гибридизация ящериц, в том числе и представителей рода Lacerta, наблюдается в природе довольно часто. Сведения об этом имеются в сводках Мертенса (Mertens, 1950, 1956, 1964), а также в более поздних работах Цвейфеля (Zweifel, 1962), Купера (Cooper, 1965а) и др. Естественные межвидовые и внутривидовые гибриды обычны и у ящериц рассматриваемой нами группы, причем наиболее подробно изучены случаи гибридизации между некоторыми обоеполыми и партеногенетическими видами в полосе соприкосновения их ареалов. Как правило, такие зоны гибридизации образуются в ущельях горных рек, по которым обоеполые формы как бы вклиниваются в область распространения партеногенетических видов. В результате возникающей здесь спонтанной гибридизации появляются гибриды, составляющие местами от 5 до 8% особей смешанной популяции. Более других нами были изучены гибриды между партеногенетическими самками L. armeniaca и самцами L. s. valentini, встречающиеся в узкой гибридной зоне на юго-западном берегу оз. Севан в Армении. Гибридные особи выводились также в лаборатории при инкубации яиц оплодотворенных самок L. armeniaca, отлавливаемых в природе приблизительно за 1-2 недели до наступления кладки или спаривавшихся при содержании их совместно с самцами в вольере. Свидетельством имевшего место спаривания в каждом отдельном случае служили остающиеся на брюхе самки четкие следы челюстей удерживавшего ее самца (рис. 75). Если отдельные партеногенетические самки, судя по наличию с каждой стороны ряда таких следов, спаривались неоднократно, то у других следы эти были единичны или же отсутствовали вовсе. Последнее обстоятельство можно объяснить тем, что при спаривании самцы реагируют прежде всего на самок, окраска которых соответствует определенному стереотипу, характерному для данной формы. Так, у кавказских скальных ящериц окраска брюха варьирует от матово-белой до розовой, оранжево-красной, оранжевой, желтой и зеленой (табл. I, II). Что же касается L. armeniaca и L. s. valentini, то у первой брюхо обычно яично-желтое, а у второй зеленовато-желтое или бледно-лимонно-желтое. Вместе с тем встречаются отдельные самки armeniaca, окраска которых по яркости приближается к таковой у отцовского вида. При содержании в вольере выяснилось, что чаще других спаривались самки, обладающие именно такой, относительно яркой окраской брюха. В этой связи следует отметить, что в смешанных популяциях скальных ящериц самцы при спаривании всегда интенсивно разыскивают именно "своих" самок. Например, в исследованной нами в северной Армении (окр. Степанавана) гибридной зоне самки бисексуальной формы L. s. portschinskii встречаются очень редко, примерно одна на 50 самок L. armeniaca. Тем не менее в июне каждая из них имела на брюхе оставшиеся после спаривания многочисленные следы челюстей, тогда как у обитающих здесь же партеногенетических самок следы эти обнаруживались крайне редко.
По своему внешнему виду рассматриваемые гибриды четко отличаются от каждой из родительских форм, и их гибридная природа легко устанавливается по совокупности ряда морфологических признаков. Ранее нами было показано, что по характеру окраски и основным показателям фолидоза они ближе стоят к партеногенетической материнской форме, нежели к бисексуальной отцовской. Вместе с тем некоторые гибридные признаки сложились у них явно под отцовским влиянием (Даревский и Куликова, 1962).
Подобную же картину дают морфологические признаки исследованных нами гибридов между партеногенетическими самками L. dahli и самцами L. s. portschinskii (Даревский и Куликова, 1961). Среди прочих гибридных признаков особенно обращают на себя внимание некоторые специфические особенности фолидоза, в частности сближение центральновисочного и барабанного, между которыми появляется обычно один сравнительно крупный промежуточный щиток (рис. 80).
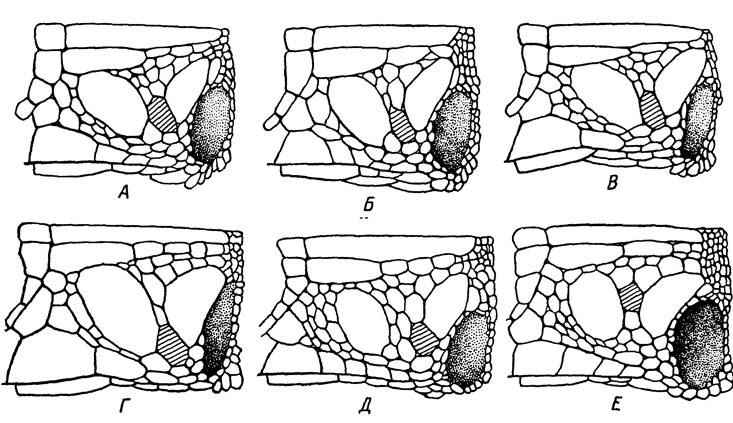
Рис. 80. Характерные особенности фолидоза височной области у гибридов между партеногенетическими и бисексуальными ящерицами: А, Б - L. armeniaca X L. s. valentini; В - L. armeniaca X L. r. macromaculata; Г - L. armeniaca X L. r. obscura; Д - L. unisexualisX L. s. valentini; E - L. s. parvula X L. derjugini. Крупный промежуточный щиток заштрихован
Ранее нами было показано, что рассматриваемые гибриды во всех случаях являются стерильными триплоидными самками (3n=57), бесплодие которых обусловлено глубокими аномалиями в развитии и строении гонад (Даревский и Куликова, 1964; Даревский, 19666). Появление их всегда наблюдается в случаях (табл. 39), когда в результате частичного перекрывания ареалов партеногенетические самки получают возможность спариваться с обитающими совместно самцами соседних бисексуальных форм.

Таблица 39. Родительские пары партеногенетических и бисексуальных ящериц, между которыми известны естественные триплоидные гибриды
Обусловленные полиплоидией крупные размеры тела таких гибридов, так же как наблюдающиеся у них четкие морфологические отличия, дают формальное право систематику, оперирующему с мертвым коллекционным материалом, рассматривать их на правах хорошо отличающихся видов (фот. 27).

Фот. 27. Схема естественной гибридизации партеногенетических самок скальных ящериц с самцами бисексуальных форм. А - L. armeniaca (самка); Б - L. s. valentini (самец); В - стерильный триплоидный гибрид; Г - L. unisexualis (самка); Д - L. s. valentini (самец); Е - стерильный триплоидный гибрид
Интересно, что подобная ошибка была допущена в свое время при описании североамериканского вида ящерицы Cnemidophorus perplexus Baird, являющегося, как теперь выясняется, лишь триплоидным гибридом между партеногенетическим видом С. neomexicanus и обоеполым С. inornatus (Lowe a. Wright, 1966). Однако полная стерильность гибридных особей превращает их в своеобразный эволюционный тупик, не имеющий значения для процесса формообразования в группе скальных ящериц.
Как показывают наши исследования, частые случаи гибридизации скальных ящериц имеют место также и в зонах совмещения ареалов соседних обоеполых форм. Хорошо выраженные гибридные зоны такого рода существуют на Северном Кавказе между L. s. saxicola и L. s. darevskii, а также между L. s. darevskii и L. s. brauneri (рис. 1). В Закавказье аналогичное положение наблюдается на стыках ареалов между L. s. portschinskii и L. s. raddei и между L. r. obscura и L. r. macromaculata.
Например, состоящая из 13 особей выборка из ущелья нижнего течения р. Гетика в северной Армении (ЗИН, № 17841) содержит 5 ящериц, являющихся чистыми L. s. portschinskii, 4 ящерицы L. s. raddei а остальные - гибриды, сочетающие признаки обеих родительских форм. Образующиеся в таких случаях межподвидовые гибриды вполне плодовиты и узнаются обычно по характерным смешанным признакам фолидоза и окраски. Именно характеризующий их широкий размах фенотипической изменчивости позволяет отличать гибридов от так называемых промежуточных особей, существование которых в смешанных популяциях обусловлено процессом первичной интерградации.
Выше (стр. 179) мы уже говорили, что между таким полусимпатрическими видами, как L. rudis и L. caucasica или L. rudis и L. saxicola, межвидовая гибридизация практически не происходит благодаря действию биологических изолирующих механизмов. Можно полагать, что именно их существованием объясняется и малая результативность опытов по скрещиванию разных видов стенных ящериц, поставленных в свое время Лантцем (Lantz, 1926).
Однако, с другой стороны, известны случаи, когда между пограничными видами скальных ящериц существует широкая зона вторичной интерградации, обусловленная практически неограниченной гибридизацией. Подобное явление очень ярко выражено, например, в зоне контакта высокогорной формы L. с. caucasica и обитающим в предгорьях подвидом L. s. daghestanica, ареалы которых отчасти перекрываются в горном Дагестане и северо-восточном Азербайджане (рис. 64). Наряду с хорошо выраженными особями каждого из симпатриантов в пограничной зоне всегда многочисленны образующиеся в разных сочетаниях гибриды, и фактически вся смешанная популяция нечистокровна. Так, например, среди 52 экземпляров ящериц из окр. Рутула в Дагестане (ЗИН, № 17732) 16 особей, или 30.7%, определяются как более или менее чистые L. s. daghestanica, И особей, 21.1%, - как L. caucasica, тогда как остальные обладают в той или иной мере выраженными гибридными признаками и точному определению не поддаются. Неоднородность указанной выборки хорошо видна также на графике (рис. 49). Подобная же картина обнаруживается и в выборках из других точек сопряженного ареала этих видов на территории Дагестана, горной Чечено-Ингушетии и южной Осетии. Аналогичная зона вторичной интерградации наблюдается в смешанной гибридной популяции L. r. rudis и L. s. lantzicyreni на Черноморском побережье северо-восточной Турции (стр. 103). В каждом из этих случаев мы сталкиваемся с положением, хорошо охарактеризованным Н. И. Лариной (1961), когда гибридизация симпатрических видов приводит к тому, что на значительном участке ареала морфологические границы симпатриантов становятся неясными и определение видовой принадлежности членов гибридной популяции весьма затруднительно. Особый интерес представляет случай, когда такая спонтанная гибридизация происходит между каким-либо из подвидов L. saxicola и экологически отличным лесным видом - артвинской ящерицей (L. derjugini Nik.). Хотя оба эти симпатрических вида обладают совершенно различными биотопами (лесная L. derjugini никогда не заходит на скалы), у подножия скальных местообитаний в лесах между ними нередко возникают контакты, приводящие к перекрестному спариванию и возникновению более или менее обширных гибридных зон. В результате происходящей затем вторичной гибридизации между гибридами и родительскими формами ящерицы из смешанных популяций отличаются необычайно высокой изменчивостью, проявляющейся в непостоянстве окраски и рисунка, а также основных признаков фолидоза. В ряде случаев при этом происходит почти полное растворение одной или обеих родительских форм в массе гибридных особей, как это наблюдается, например, в Боржомском ущелье и в окр. Абастумани в зоне гибридизации между L. derjugini и L. s. parvula. Одна из таких гибридных зон в окр. Бакуриани в Грузии была изучена недавно В. Ф. Положихиной (1965). Возможность естественной гибридизации между L. s. parvula и L. derjugini была проверена нами также путем их реципрокного скрещивания в эксперименте. Постановка этого опыта определялась тем обстоятельством, что одна из форм скальной ящерицы, именно L. mixta, как уже говорилось выше (стр. 140), была описана в качестве вероятного гибрида между L. saxicola и L. derjugini (Mehely, 1909).
В результате ряда экспериментов, поставленных на базе Зоологического института АН АрмССР в Ереване, нами было получено 7 межвидовых гибридов от самок L. derjugini, оплодотворенных при вольерном содержании самцами L. s. parvula, и 2 гибрида от обратного скрещивания в природных условиях. В последнем случае отловленные ранней весной в окр. Абастумани неоплодотворенные самки L. s. parvula были выпущены в окр. Бакуриани на изолированные посреди леса вдали от скал каменные развалины, заселенные исключительно L. derjugini, Изучение полученных гибридов показало, что по ряду признаков фолидоза, в частности расположению височной области, большинство из них мало отличается от типичных L. mixta из окр. Абастумани. Отметим, что характерное для последней формы и наблюдающееся также у гибридов укрупнение промежуточного щитка между центральновисочным и барабанным свойственно также и другим гибридам ящериц рассматриваемой группы (рис. 80, Е).
Таким образом, гибридная природа L. mixta действительно очень вероятна, хотя скорее всего эта форма представляет собой не первое поколение гибридов, а выкристаллизовывается постепенно в результате неоднократных последующих скрещиваний "в себе". В данной связи можно отметить, что в популяциях L. mixta часто встречаются особи, по окраске и рисунку тела практически не отличимые от типичных L. derjugini (фот. 20, Б, Ж). Интересно также, что наблюдаемые в природе гибриды сравнительно редко встречаются на скалах, а держатся больше у их подножия в лесу, т. е. обнаруживают биотопические требования, сходные с таковыми у L. derjugini.
Известно, что мнение о положительной роли гибридизации как факторе формообразования у животных имеет своих противников и сторонников и неоднократно обсуждалось в литературе. Критическое рассмотрение этого вопроса позволило С. С. Шварцу (1959) аргументировать точку зрения, что межвидовая гибридизация даже в случае, если она представляет собой закономерное явление, не может играть важной роли в эволюционном процессе. Вместе с тем им не отрицаются и отдельные,, приводимые в литературе случаи гибридного происхождения некоторых видов позвоночных. К последним можно добавить также интересную работу Сандерса (Sanders, 1961), указывающего на возможность гибридного происхождения североамериканской жабы Bufo terrestris. Положительная формообразующая роль гибридизации была обстоятельно аргументирована недавно Н. П. Лариной (1961), по мнению которой существование стабилизировавшихся гибридных популяций у позвоночных животных позволяет считать межвидовую гибридизацию одной из вероятных причин и одним из источников формообразований. Рассмотренные нами случаи спонтанной гибридизации у ящериц подрода Archaeolacerta также свидетельствуют в пользу такой точки зрения.
Особый интерес представляет рассмотренный нами ранее вопрос о положительной роли гибридизации в становлении партеногенетических видов скальных ящериц (Даревский, 1962; Darevsky, 19666). Отметим, что, по данным Лоу и Райта (Lowe a. Wright, 1966а, 19666), гибридным происхождением обладают также некоторые партеногенетические виды ящериц рода Cnemidophorus.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'