Происхождение и филогения кавказских скальных ящериц подрода Archaeolacerta
Едва ли следует сомневаться в том, что первичным центром происхождения политипической группы ящериц подродов Podarcis и Archaeolacerta служила территория Южной и Юго-восточной Европы, объединяемая обычно под общим названием восточноевропейского Средиземноморья. Это доказывается, в частности, тем, что в настоящее время в пределах указанной территории обитают 19 из 27 известных видов обоих подродов, многие из которых в свою очередь распадаются на значительное число подвидов. Так, согласно последнему каталогу европейской герпетофауны (Mertens u. Wermuth, 1960), такие широко распространенные виды, как Lacerta erhardii, L. pityusensis, L. sicula и L. muralis, образуют здесь соответственно 31, 32, 39 и 18 четко различающихся подвидовых форм, причем такой же полиморфизм свойствен и большинству других европейских представителей рассматриваемой группы. Хотя многие из этих подвидов имеют сравнительно недавнее - четвертичное происхождение (Kramer u. Mertens, 1938; Eisentraut, 1849; Wettstein, 1953; Radovanovic, 1959), представляется вероятным, что их исходные формы образовались здесь гораздо раньше - еще в третичное время, учитывая, в частности, тот факт, что представители собственно рода Lacerta известны из Южной Европы (Франция) уже с эоцена. Можно отметить, что к аналогичному выводу в отношении группы собственно L. muralis уже пришли ранее Дехо и Ландон (Dchaut et Landon, 1950), по мнению которых центром происхождения этого полиморфного вида является южное Средиземноморье, в частности территория современной Италии и Сицилии.
Попытаемся представить себе, в каких условиях должно было протекать формирование исходного экологического типа ящериц, от которых произошли затем современные представители подродов Podarcis и Archaeolacerta. Согласно имеющимся палеогеографическим данным, еще в миоцене Южная Европа выглядела не сплошной материковой сушей, а представляла собой архипелаги более или менее сближенных островов. Гористые острова располагались, в частности, на месте современной Италии, в то время как Корсика, Сардиния и некоторые другие острова Тирренского моря образовывали единую сушу. Что касается Балканского полуострова, то в миоцене он представлял собой уже довольно обширную сушу, окружавшую древнее Родопское поднятие, причем в этот период Балканы являлись полуостровом Западной Азии, а не Европы (Вульф, 1944). Западная часть Греции, так же как юг Малой Азии, и нижний Египет в это время были покрыты морем, тогда как восточная часть Пелопонесского полуострова образовывала единое целое с Малой Азией, будучи связана с ней сушей, располагавшейся на месте современного Эгейского моря. Лишь в плиоцене произошло опускание эгейской суши, на месте которой образовались современные Мраморное и Эгейское моря с их многочисленными островами. При этом долины древних рек Геллеспонта и Босфора превратились в проливы, соединяющие Черное и Средиземное моря, и балканская суша, потеряв свою связь с Азией, на всем протяжении соединилась с Европой, превратившись в полуостров последней.
По данным Е. Н. Вульфа (1944), в начале третичного периода лесная флора Южной Европы носила ярко выраженный тропический характер, хотя здесь в ограниченном количестве имелись и древесные виды с опадающей листвой, представляющие собой зачатки будущей флоры умеренного типа. Важно отметить, что в составе ее уже тогда существовали отдельные роды и виды, входящие в комплекс современных средиземноморских растительных группировок, что "не оставляет никакого сомнения в преемственности этих флор и дифференциации средиземноморской флоры из состава тропической флоры" (Вульф, 1944, стр. 137). Можно, таким образом, полагать, что формирование исходных форм интересующей нас группы ящериц протекало в условиях тропического леса, покрывавшего в третичное время склоны отдельных хребтов и гор, причем наличие на рассматриваемой территории многочисленных изолированных морем участков суши с самого начала должно было обусловить возникновение характерного для подрода Podarcis полиморфизма.
По всей видимости, первоначально ящерицы рассматриваемой группы эволюционировали по пути приспособления к полудревесному образу жизни, в частности к обитанию на стволах деревьев, как это имеет место у некоторых современных лесных видов, например у распространенной в юго-восточном Азербайджане Lacerta chlorogaster. Переход же их к жизни на поверхности открытых скал, т. е. превращение из древесных в "стенных" и "скальных", произошел уже вторично и не потребовал выработки каких-либо особых дополнительных экологических адаптаций. Благодаря неоднократно устанавливавшейся в третичное и четвертичное время сухопутной связи Юго-восточной Европы с Малой Азией стенные ящерицы получили возможность расселения на восток до Кавказа включительно, где, как мы покажем дальше, образовался вторичный очаг формообразования, приведший к возникновению многочисленных кавказских представителей подрода Archaeolacerta.
Таким образом, мы полагаем, что группа скальных ящериц этого подрода филогенетически более молода, нежели группа собственно стенных ящериц подрода Podarcis, разделяя в этом отношении точку зрения, ранее уже высказанную К. Клеммером (Klemmer, 1957). Отметим, что этот исследователь пришел к аналогичному выводу на основании анализа ряда краниологических признаков, справедливо считая, в частности, что характерное для большинства Podarcis наличие птеригоидных зубов в сочетании с большим окостенением надглазничной области крыши черепа является свидетельством их значительной древности. Косвенным свидетельством сравнительной молодости кавказских скальных ящериц может служить наблюдающееся у гибридных форм внутри этой группы проявление некоторых анцестральных признаков, в норме характерных для европейских представителей подродов Podarcis и Archaeolacerta.
Мы уже говорили (стр. 190), что спорадически встречающаяся в западной Грузии высокогорная ящерица Lacerta mixta исходно возникла благодаря естественной гибридизации между L. saxicola и лесным видом L. derjugini. Хотя обе эти родительских формы характеризуются одноцветными, без пятен, брюхом и нижней стороной головы, в популяциях L. mixta изредка встречаются особи с явственной пятнистостью брюха, в норме свойственной таким южноевропейским видам, как L. muralis, L. bedriagae, L. danfordi и др. Точно так же обычное у гибридов между бисексуальными и партеногенетическими видами скальных ящериц укрупнение щитков височной области нормально наблюдается у балканской L. horvathi, причем последний вид характеризуется также типом окраски и рисунка, характерным для упоминавшейся выше L. mixta.
Как анцестрапьные признаки могут рассматриваться и некоторые своеобразные особенности фолидоза, наблюдающиеся у всех партеногенетических видов, возникновение которых, как мы полагаем, исходно связано с гибридизацией. Так, характерное для L. dahli частое удвоение задненосового щитка обычно у южноевропейских видов L. oxycephala, L. danfordi и др., а свойственное армянской L. unisexualis разделение межчелюстного и лобоносового щитков (рис. 59, А) служит характерным отличительным признаком балканского вида L. horvathi. Отметим, что проявление в гибридном потомстве предковых признаков неоднократно описывалось также у ряда птиц, млекопитающих, рыб и беспозвоночных, причем такая атавизация наблюдается и при скрещивании подвидов и пород в пределах одного вида (Шварц, 1959).
Обращает на себя внимание значительное сходство отдельных форм скальных ящериц с некоторыми европейскими представителями того же подрода. Например, распространенная в южном Закавказье L. s. valentini по внешнему виду чрезвычайно напоминает пиренейскую L. monticola, а живущая на Большом хребте L. caucasica обладает значительным сходством с балканской L. horvathi. Интересно, что аналогичные примеры известны и энтомологам, выдвинувшим для их объяснения некоторые зоогеографические соображения. Так, Жаннель (цит. по: Яблоков-Хизорян, 1961), отмечая значительное габитуальное сходство кавказского и пиренейского подродов жужелиц (Cechenus Zish и Iniopachys Sol.), склонен рассматривать их как реликты палеогеновой фауны, изолировавшиеся во время морской трансгрессии олигоцена на крайних точках их первоначального ареала. Подобная гипотеза действительно выглядит подкупающей, однако, как справедливо замечает по этому поводу С. М. Яблоков-Хизорян (1961), геологами не было обнаружено никаких намеков на существование олигоценовой континентальной связи между Пиренеями и Кавказом. Что же касается рассмотренного выше примера с ящерицами, то можно полагать, что здесь мы имеем дело со случаями конвергентного сходства, обусловленного параллельной эволюцией в сходных условиях существования. Подобное предположение хорошо согласуется с точкой зрения К. Клеммера (Klemmer, 1957), высказавшегося в пользу полифилитического происхождения ящериц группы Archaeolacerta. В свете такого допущения может быть понято и своеобразие современного ареала этого подрода (рис. 81), отдельные виды которого встречаются вне связи друг с другом на Пиренейском полуострове (L. monticola), в Сардинии и Корсике (L. bedriagae), северо-западной Югославии (L. horvathi, L. oxycephala), южной Греции (L. danfordi), Малой Азии, в Крыму и на Кавказе (L. saxicola). Для решения вопроса, каким образом протекала эволюция рассматриваемых нами ящериц именно в пределах Кавказа, необходимо хотя бы в первом приближении выяснить, какие из многочисленных современных форм этой группы филогенетически более древние. Рассуждая теоретически, такие древние формы следует искать прежде всего в убежищах третичных реликтов флоры и фауны, не пострадавших в эпоху четвертичных оледенений. Как мы уже отмечали, по данным ботаников (Буш, 1935; Гроссгейм, 1948), в пределах Кавказа существовало несколько подобных убежищ, наиболее значительные из которых располагались на территории современной Колхиды и в Талыше (убежища мезофильной растительности) и в южной Армении и Дагестане (убежища ксерофильных растительных группировок). Более мелкие рефугиумы теплолюбивой фауны существовали и в некоторых других районах Кавказского перешейка, в частности в долине Куры. Отбирая по данному принципу предположительно наиболее древних ящериц интересующей нас группы, мы получим следующий ряд форм, основные современные ареалы которых располагаются в границах названных выше реликтовых территорий.
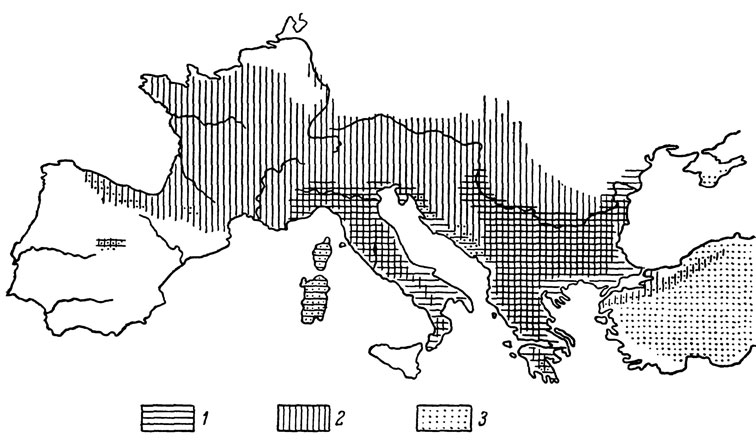
Рис. 81. Современное распространение в Европе и Малой Азии скальных ящериц подродов Podarcis и Archaeolacerta. (По К. Клеммеру, 1957, с дополнениями): 1, 2 - общий ареал видов подрода Podarcis; 2 - ареал L. muralis; 3 - общий ареал подрода Archaeolacerta
Можно видеть, что отобранные таким образом в качестве предположительно наиболее древних формы ящериц габитуально весьма отличаются друг от друга, однако легко показать, что все они обладают рядом общих морфологических признаков, касающихся прежде всего некоторых меристических особенностей чешуйчатого покрова. Указанное обстоятельство является особенно важным и на нем необходимо остановиться подробнее. Дело в том, что некоторыми систематиками уже давно было показано, что уменьшение количества элементов чешуйчатого покрова, сопровождающееся, как правило, увеличением размеров отдельных щитков и чешуй, является эволютивным признаком, довольно четко прослеживающимся в филогенезе отдельных форм. В частности, у ящериц подобная зависимость была установлена Вермутом (Wermuth, 1955) и Стугреном и Ванча (Stugren si Vancea, 1961) для L. vivipara и Стугреном (Stugren, 1962) - для L. agilis. Отмеченная этими авторами закономерность является лишь частным случаем универсального явления, известного под названием "закона олигомеризации гомологичных органов", впервые сформулированного В. А. Догелем (1954). Одно из основных положений этого закона заключается, по В. А. Догелю, в том, что эволюция различных групп Metazoa сопровождается постепенным уменьшением числа гомологичных органов, чему очень часто сопутствует увеличение их размеров и интенсификация функций. Среди многочисленных примеров, иллюстрирующих этот закон применительно к различным группам животных, В. А. Догель приводит и пресмыкающихся, отмечая в первую очередь четко прослеживающуюся в их эволюции олигомеризацию первичного рогового покрова из мелких роговых зернышек в более крупные метамерные образования. Рассматривая в свете этой закономерности эволюцию кавказских ящериц подрода Archaeolacerta, мы имеем все основания считать, что она сопровождалась увеличением размеров и, следовательно, сокращением числа отдельных элементов чешуйчатого покрова, характеристика которого имеет решающее значение для систематики. Иными словами, формы с максимальным числом таких элементов должны рассматриваться как филогенетически более древние.
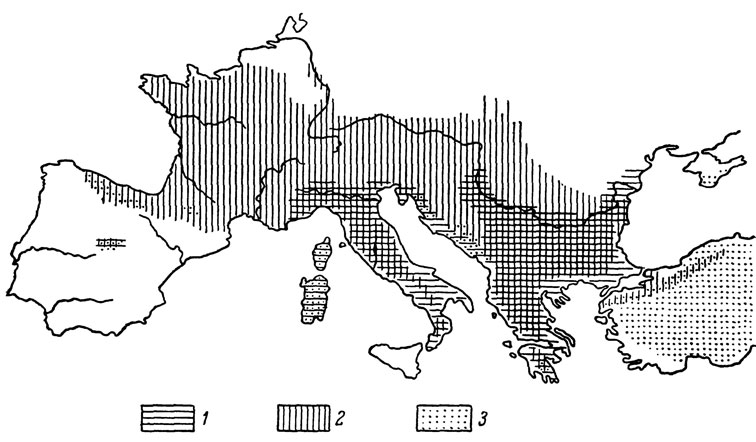
Рис. 82. Сравнительное количество чешуи вокруг середины тела у различных кавказских форм скальных ящериц: 1 - L. s. szczerbaki; 2 - L. s. darevskii; 3 - L. s. lindholmi; 4 - L. s. brauneri; 5 - L. s. parvula; 6 - L. s. nairensis; 7 - L. s. saxicola; 8 - L. s. raddei; 9 - L. s. daghestanica; 10 - L. s. valentini; 11 - L. r. obscura; 12 - L. r. macromaculata; 13 - L. r. rudis; 14 - L. c. alpina; 15 - L. c. caucasica
Указанная закономерность хорошо подтверждается прежде всего на таких важных диагностических признаках, как число туловищных чешуй в одном ряду вокруг середины тела и размеры щитков височной области. Располагая по этим признакам всех кавказских представителей подрода Archaeolacerta, их можно разделить на две хорошо различающиеся группы, объединяющие "многочешуйные" формы, с одной стороны, "малочешуйные" - с другой. Как видно на рис. 82 и 83, первую из них составляют именно перечисленные выше подвиды L. saxicola, сохранившиеся в убежищах третичных реликтов. Другой важный вывод, вытекающий из сделанного сопоставления, заключается в том, что интенсивная эволюция скальных ящериц, приведшая к возникновению ряда "малочешуйных" форм, протекала в пределах Кавказа уже в послеледниковое время. Один из путей такого формообразования связан, по нашему мнению, с процессом расселения ящериц в горы вслед за отступающим ледником. Выше мы уже говорили, что в смешанных популяциях скальных ящериц в полосе соприкосновения ареалов пограничных форм осуществляется механизм репродуктивной изоляции, основанный на разновременности сроков созревания гонад. Высокогорные формы при этом приступают к размножению, как правило, значительно раньше распространенных в предгорьях, что биологически вполне оправдано. Легко допустить, что подобное ускорение сроков размножения могло выработаться у расселяющейся в горы популяции внутри одного вида, как это наблюдается у некоторых широко распространенных современных форм, например у L. s. portschinskii в Армении и Грузии. В таком случае вновь образовавшаяся горная часть популяции окажется вскоре репродуктивно изолированной на периферии ареала со всеми вытекающими отсюда последствиями. Именно подобным дивергентным путем образовался, по нашему мнению, высокогорный вид L. caucasica, отделившийся в послеледниковое время от L. s. daghestanica, что подтверждается явной преемственностью их морфологических признаков. При возможном вторичном смешении популяций исходной и вновь образовавшейся формы возникшее расхождение в сроках созревания гонад может остаться и обе они сохранят свою самостоятельность, несмотря на наличие полусимпатрических ареалов.
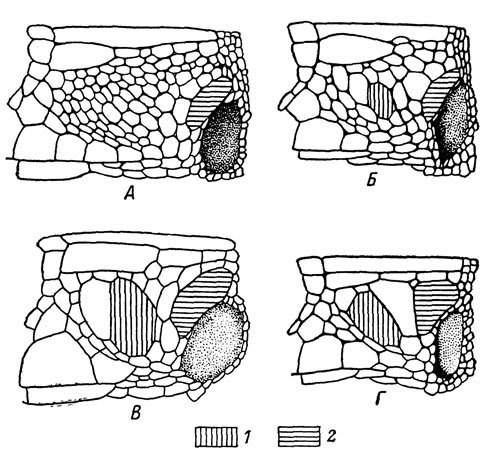
Рис. 83. Последовательное укрупнение элементов чешуйчатого покрова височной области: А - L. s. szczerbakii; Б - L. с. alpina; В - L. armeniaca; Г - L. mixta. Щитки: 1 - центральновисочный, 2 - барабанный
Между прочим, хорошим доказательством послеледникового происхождения этого вида является отсутствие его в западном Закавказье (Месхетский хребет), где сохранился другой ледниковый реликт - прометеева полевка (Prometheomys schaposchnikovi), расселившаяся сюда с Главного Кавказского хребта в плейстоцене (Верещагин, 1959).
О формообразующей роли естественной гибридизации в группе Archaeolacerta мы уже говорили выше. Ранее было также показано, что возникновение партеногенетических видов скальных ящериц от исходных бисексуальных форм произошло в сохранившихся лесных рефугиумах, видимо, в эпоху четвертичных оледенений Кавказа (Даревский и Куликова, 1961; Даревский, 1962, 1966а).
Учитывая все вышеизложенное, мы с достаточной долей вероятности можем следующим образом представить себе пути эволюции и расселения скальных ящериц в пределах Кавказа.
По всей видимости, в миоцене из Малой Азии на Кавказ впервые проникают, скорее всего сразу несколько, исходные лесные формы ящериц группы Podarcis-Archaeolacerta. Одним из таких древних вселенцев является сохранившийся в лесах Талыша до наших дней лесной реликтовый вид - Lacerta chlorogaster. Дальнейшее расселение их на Кавказе проходило вдоль Черноморского побережья, с одной стороны, и в обход Малого Кавказа через долины Аракса и Куры - с другой. Не исключен также и дальнейший путь в обход Главного хребта с востока, что подтверждается существованием в Дагестане многих закавказских фаунистических элементов, в частности ящериц и змей. Расселяющаяся вдоль побережья западная ветвь малоазиатских вселенцев достигла северо-западной оконечности Большого хребта в районе современной Анапы, где до настоящего времени сохранилась изолированная реликтовая популяция древней "многочешуйной" формы L. s. szczerbaki. К последней близко примыкает отделившаяся от нее более молодая (судя по уменьшению числа элементов чешуйчатого покрова) и более влаголюбивая L. s. darevskii, давшая затем два близких подвида - L. s. brauneri и L. s. saxicola. Вероятно, в начале плейстоцена от этой группы отделилась и крымская форма L. s. lindholmi, что произошло в период существования кратковременной перемычки суши между Крымским полуостровом и Кавказом. Интересно отметить, что крымская форма обнаруживает значительное сходство не с территориально более близкой L. s. szczerbaki, а с распространенной юяшее по побережью L. s. darevskii. Это обстоятельство позволило Н. Н. Щербаку (1962а) высказать в свое время справедливое мнение, что сухопутная связь между Кавказом и Крымом существовала где-то в районе современных ареалов этих подвидов, а не по кратчайшей линии между Таманским и Керченским полуостровами.
Вероятно, независимо от рассмотренных выше форм в западное Закавказье проникла позднее современная L. s. parvula. Что касается восточной ветви малоазиатских вселенцев, то дальнейшая эволюция их в пределах Кавказа представляется нам следующим образом. Одной из первых в древнюю долину Куры проникает адаптировавшаяся к сухим местообитаниям L. s. portschinskii, от которой путем перехода к партеногенезу в ледниковое время произошла морфологически очень близкая к ней, также сухолюбивая L. rostombekovi.
Группу тесно связанных в морфологическом отношении форм образуют в восточном Закавказье и северном Иране L. s. nairensis, L. s. raddei и L. s. defilippii, от первой из которых отделилась позднее партеногенетическая L. unisexualis, тогда как от второй аналогичным образом произошла очень близкая к ней L. dahli. Наконец, уже в послеледниковое время из северо-восточной Турции, возможно, вторично расселяется в Закавказье высокогорная L. s. valentini, по некоторым признакам близкая к партеногенетическому виду L. armeniaca.
Особый интерес представляет вопрос о происхождении дагестанского подвида L. s. daghestanica. Имеются основания полагать, что эта ящерица проникла в Предкавказье вместе с другими закавказскими фаунистическими элементами, расселяясь с юга вдоль берегов древнего Куринского залива, существовавшего еще в конце миоцена (Хайн и Шарданов, 1952). Свидетельством такого миграционного пути являются, в частности, реликтовые популяции этого подвида, сохранившиеся на юго-восточных склонах Главного хребта в Нухинском и Варташенском районах Азербайджана (рис. 64). Лишь в послеледниковое время от L. s. daghestanica отделилась высокогорная форма L. caucasica, разделившаяся затем на два отдельных подвида. Можно, таким образом, видеть, что вид Lacerta saxicola распадается в пределах Кавказа на широко разобщенные географически группы подвидов - восточную и западную, общность происхождения которых подтверждается также присущей тем и другим сходной манерой спаривания, подробно рассмотренной нами выше. В частности, обращает на себя внимание сходный тип удерживания самок за ногу, наблюдающийся у восточнозакавказских подвидов L. s. nairensis и L. s. raddei, с одной стороны, и у предкавказских L. s. saxicola, L. s. brauneri, L. s. szczerbaki - с другой.
Это обстоятельство хорошо увязывается с нашими представлениями о путях проникновения на Кавказ ящериц подрода Archaeolacerta, расселявшихся двумя независимыми путями по черноморскому побережью на западе и в обход Малого Кавказа с юго-востока. По всей видимости, исходные лесные формы этой группы обладали общей для них манерой спаривания с удержанием самки за ногу, о чем свидетельствует сохранение ее у современного реликтового лесного вида L. chlorogaster. В настоящее время этот исходный тип поведения без изменения сохранился также у восточнозакавказской L. s. nairensis, тогда как остальные формы, в частности вся группа западнокавказских вселенцев, вторично приобрела манеру удержания самки за края тела, о чем свидетельствует и поныне наблюдающийся у них смешанный тип спаривания. Таким образом, значительная морфологическая близость у географически разобщенных восточных и западных подвидов Lacerta saxicola находит свое объяснение в общности их происхождения, доказываемой на основании сходного поведения при спаривании. Морфологически хорошо выраженную группу форм образуют на Кавказе все три подвида Lacerta rudis, родственные связи которых с другими скальными ящерицами, однако, не вполне ясны. Наиболее примитивной из них несомненно является обитающая в долине Куры L. г. obscura, характеризующаяся максимальным количеством туловищных и височных чешуй. От нее произошли позднее подвиды macromaculata и rudis, последний из которых расселился затем на обширной территории Большого Кавказа и Закавказья. Не исключено, что L. s. obscura проникла в долину среднего течения Куры еще в миоцене с запада, в период существования пролива, соединявшего Куринский и Рионский бассейны. При этом допущении становится понятным и некоторое морфологическое сходство L. г. obscura с малоазиатскими формами L. s. tristis и L. s. lantzicyreni - обстоятельство, на которое в свое время уже обратили внимание Лантц и Цирен (Lantz et Cyren, 1936).
Изложенные выше соображения о филогенетических связях скальных ящериц в пределах Кавказа наглядно представлены на рис. 84.
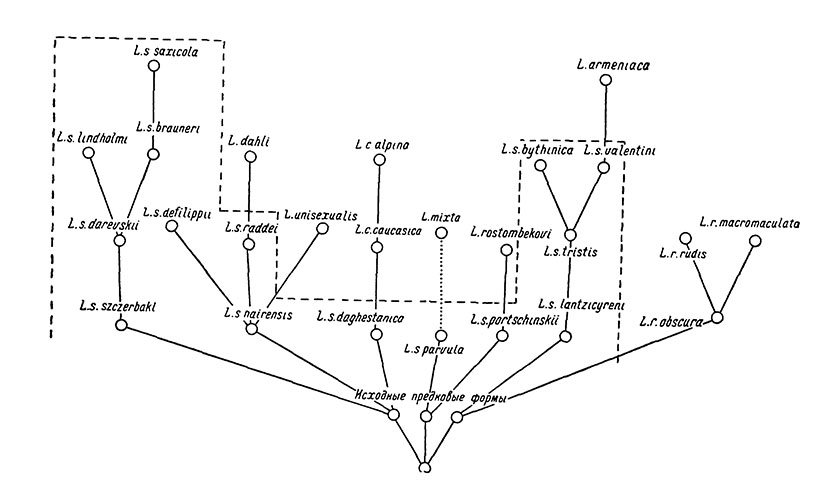
Рис. 84. Схема филогенетических отношений скальных ящериц. И сходный полиморфный вид Lacerta saxicola ограничен пунктиром; гибридогенный монотипический вид Lacerta mixta условно показан производным от одной из возможных родительских форм - L. s. parvula
Отметим, что предложенная здесь филогенетическая схема основана именно на предположительно родственных связях отдельных форм, т. е. отражает, по нашему мнению, относительную их близость к общему ближайшему предку в том смысле, как это понимает, в частности, И. И. Соколов (1965).
Огромное влияние на современное распространение скальных ящериц в пределах Кавказа, как, впрочем, и всех остальных обитающих здесь горных животных, сыграла эпоха четвертичных оледенений и связанных с нею изменений климата в сторону похолодания. Нужно подчеркнуть,, что, несмотря на большое число специальных исследований, вопрос о ледниковом периоде в пределах Кавказа все еще остается запутанным и до конца не решенным (Гвоздецкий, 1954; Марков, 1965). Большинство специалистов склоняются все же к мнению, что в плейстоцене Кавказ подвергался оледенениям дважды. Обозначая эти периоды альпийскими терминами "рисс" и "вюрм", они признают синхронность альпийского и кавказского ледниковых периодов. В переводе на язык геологической истории Кавказа эти термины соответствуют хазарскому и хвалынскому ярусам.
Не касаясь специально этого вопроса, можно отметить, что, по имеющимся литературным данным, ледниковый период в пределах Кавказа характеризовался достаточно широким развитием горных ледников, что должно было привести к коренной перестройке ареалов практически всех обитающих здесь видов растений и животных. Что же касается, в частности, ящериц рассматриваемой нами группы, то льдом и снегом в этот период была занята практически вся расположенная в горах часть их современного ареала. Ареалы многих горных животных должны были в это время неизбежно сместиться книзу не столько под воздействием самого ледника, сколько под влиянием связанного с ним общего похолодания климата. Необходимо, однако, отметить, что представления старых авторов о чрезмерно суровом климате ледниковых эпох в свете более современных данных выглядят сильно преувеличенными. Так, по Л. И. Маруашвили (1956), среднегодовая температура понижалась, видимо, не более чем на 1.5-2°, тогда как осадков (в горах восточного Кавказа) выпадало не менее 1500-2000 мм. Еще раньше к аналогичному заключению пришел А. Л. Тахтаджян (1946), по мнению которого климат ледниковых эпох в Закавказье был лишь незначительно холоднее современного и изменения его выражались лишь в снижении годовой амплитуды за счет уменьшения главным образом летнего тепла.
Хотя пояса растительности в этих условиях должны были заметно сместиться книзу, климат предгорий оставался все же достаточно мягким, что позволило местами пережить похолодание таким теплолюбивым растениям, как тис, дзельква, лавровишня, самшит, лавр и некоторые другие. Особенно показателен в этом отношении тис, реликтовые рощи и отдельные деревья которого сохранились во многих местах Большого Кавказа и Закавказья. Очевидно, что, помимо двух наиболее известных в этом отношении убежищ теплолюбивых видов - Колхидской низменности и Талыша, о которых писал в свое время А. А. Гроссгейм (1948), в различных местах Кавказа имелись и другие, хотя и менее обширные, рефугиумы. В частности, в Армении наличие подобных убежищ было показано И. С. Даревским (1959) для пресмыкающихся и С. М. Яблоковым-Хизоряном (1961) для насекомых. В свете всего вышеизложенного мы с довольно значительной долей вероятности можем представить себе общую картину распространения скальных ящериц в период максимальной фазы последнего оледенения Кавказа. Очевидно, что на покрытых в значительной своей части льдом и снегом склонах Большого хребта интересующие нас ящерицы, как и вообще всякие другие пресмыкающиеся, отсутствовали. Учитывая, однако, что на южных склонах снеговая линия не опускалась ниже 2000 м, т. е. проходила примерно на 1000 м ниже, чем в настоящее время, вполне вероятно, что на обращенных к морю склонах Гагринского, Бзыбского и некоторых других хребтов, так же как и на самом побережье, существовали условия, благоприятствующие сохранению здесь отдельных популяций Lacerta saxicola. Это относится прежде всего к L. s. brauneri и L. s. darevskii, а такя^е к распространенной в районе Анапы реликтовой форме L. s. szczerbaki. Что касается северных склонов Большого хребта, то обитавшие здесь в теплую межледниковую эпоху пресмыкающиеся, в том числе и скальные ящерицы, были оттеснены, видимо, далеко в предгорья. В данной связи можно сослаться на мнение А. А. Гроссгейма (1948), считавшего, что "развитие ледниковых явлений у северного подножия Большого Кавказа послужило причиной массового вымирания здесь средиземноморцев и современной бедности этого района средиземноморскими элементами". Видимо, замечает по этому поводу Н. А. Гвоздецкий (1954), дело здесь не только в непосредственном ВЛИЯНИИ ледников, но и в том, что к северу от горного барьера Большого Кавказа климат в ледниковые эпохи становился чрезмерно суровым. Вероятно, к этому периоду следует относить и проникновение скальных ящериц на Ставропольское плато, где они до самого последнего времени существовали в соответствующих местообитаниях среди реликтовых островных лесов. С другой стороны, вызывает удивление тот факт, что ящерицы данной группы отсутствуют в очень подходящих для них условиях на склонах пятигорских локкалитов, образовавшихся, как полагают, еще в неогене. В то же время здесь обнаружен такой реликтовый вид, как снежная полевка (Microtus nivalis), несомненно расселившаяся с Большого Кавказа в ледниковый период (Верещагин, 1959). Не исключено, однако, что L. saxicola исчезла в этих местах уже в более позднее время, как произошло это уже на наших глазах в некоторых местах Ставрополья.
Значительное смещение ареалов скальных ящериц под влиянием оледенения происходило также и в Закавказье. Отдельные популяции L. s. parvula и L. rudis могли сохраниться в западном Закавказье, в частности в районе Колхидской низменности, являвшейся, как уже говорилось, одним из главных убежищ теплолюбивой флоры и фауны. По всей видимости, ящерицы этой группы обитали и на Черноморском побережье в современной Аджарии и северо-восточной Турции, поскольку на южных склонах Месхетского хребта снеговая линия не спускалась ниже 2000-2200 м. Как и в настоящее время, скальные ящерицы были, видимо, широко распространены на территории современного Талыша и Ленкоранской низменности, где влияние ледника проявилось лишь в незначительном похолодании и увлажнении климата. Развитие ледникового покрова на южных склонах Большого хребта, с одной стороны, и на северных склонах хребтов Малого Кавказа - с другой, должно было привести к оттеснению обитающих здесь животных, в том числе и пресмыкающихся, в долину Куры, также сыгравшую, видимо, немаловажную роль в качестве убежища теплолюбивой фауны. О том, что скальные ящерицы действительно были распространены здесь в тот период, свидетельствует, в частности, реликтовая популяция L. dahli, сохранившаяся благодаря выходу на поверхность холодных подземных вод в окр. сел. Карели на левом берегу Куры. Вероятно, как и в настоящее время, в долине среднего течения Куры была довольно широко распространена и L. s. portschinskii.
Глубокие изменения в распространении пресмыкающихся, как и многих других животных, несомненно имели место также на территории центрального Закавказья и, в частности, Армянского нагорья. Похолодания климата, сопровождавшиеся значительным снижением снеговой линии на южных склонах Малого Кавказа и северных склонах горы Арарат в Турции, должны были привести к отступлению теплолюбивой фауны в долину Аракса и далее к югу - на территорию современного Ирана. Что же касается обширной территории Северной Армении и южной Грузии, то имеются все основания полагать, что некоторые обитающие здесь формы скальных ящериц, в частности L. armeniaca, L. dahli и L. rostombekovi, могли пережить ледниковую эпоху в сохранившихся лесных убежищах, о которых уже говорилось выше. О наличии таких убежищ, в частности в лесной зоне северной Армении, свидетельствуют сохранившиеся здесь реликтовые рощи тиса.
Наступившее в голоцене общее потепление климата, сопровождавшееся повсеместным отступанием ледников, ознаменовало собой новый этап перестройки ареалов ящериц рассматриваемой группы. Постепенно освобождавшиеся от льда речные долины являлись, по всей видимости, наиболее удобным и естественным путем проникновения теплолюбивой фауны в горы, что в полной мере относится и к пресмыкающимся. Именно такое расселение по речным долинам обусловило в конечном счете чрезвычайно сложную и запутанную картину современных ареалов ящериц подрода Archaeolacerta в пределах Кавказа. В голоценовую эпоху на Большой Кавказ по Сурамскому хребту проникает, видимо, L. rudis, широко расселившаяся затем к востоку и западу по его южным склонам. К этому времени следует относить и возникновение эндемичных высокогорных форм.
Значительную роль в распространении ряда видов и подвидов ящериц сыграла также позднечетвертичная вулканическая деятельность, в результате которой огромные территории были залиты лавой. Применительно к некоторым закавказским формам этот вопрос рассматривался уже нами ранее (Даревский, 1959, 1960а).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'