Питание
Подобно всем остальным представителям своего семейства, скальные ящерицы преимущественно энтомофаги, состав пищи которых определяется прежде всего соотношением в биоценозе различных объектов добычи, в соответствии с чем заметно меняется по сезонам. Представление о питании некоторых закавказских форм дают таблицы 32, 33, основанные на анализах содержимого 350 желудков взрослых особей, отловленных нами совместно с Ф. Д. Даниеляном на протяжении всего сезона активности, с апреля по сентябрь.
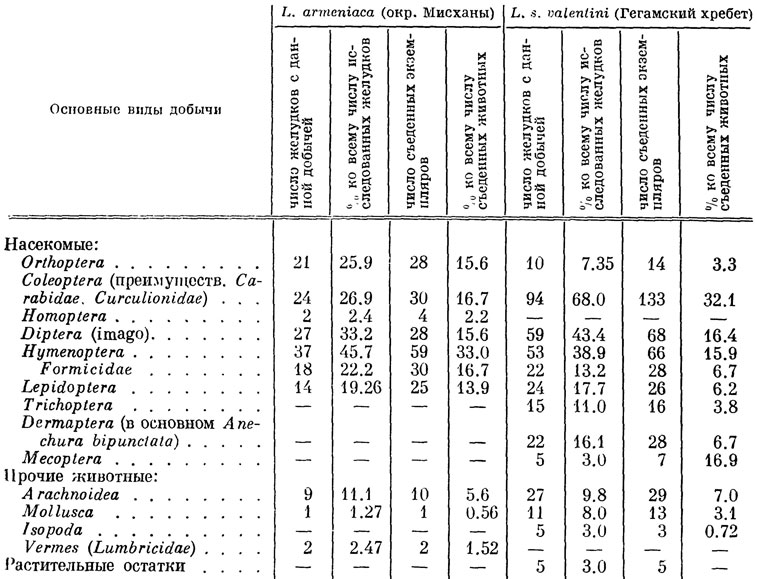
Таблица 32. Питание L. armeniaca и L. s. valentini в апреле-сентябре в условиях Армении
Обращает на себя внимание сравнительно высокий процент содержания в пище исследованных ящериц двукрылых и перепончатокрылых насекомых, среди которых встречаются многие подвижные и быстро летающие формы. Как показывают наблюдения в природе, ящерицы часто ловят их на лету, совершая стремительные и точные прыжки и безошибочна схватывая пролетающее насекомое. Кроме того, по утрам после дождливых и холодных ночей они выходят на освещенные камни и быстро нагреваются солнцем. Затем, спускаясь в траву, схватывают скованную холодом, неподвижную добычу. Из других поедаемых животных заметное место занимают саранчовые, мелкие жуки (долгоносики, жужелицы, чернотелки и листоеды), а также небольшие бабочки, гусеницы и пауки. Размеры добычи варьируют при этом от сравнительно крупных, длиной до 4 см, голых гусениц до муравьев и тлей.
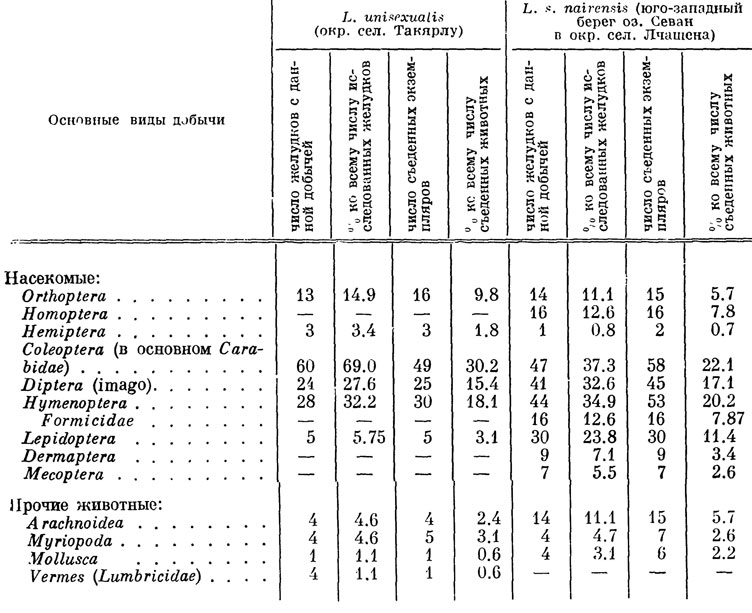
Таблица 33. Питание L. unis xualis и L. s. nairensis в апреле-сeнтябре в условиях Армении
Наличие в желудках взрослых ящериц очень мелких пищевых объектов объясняется своеобразной манерой поведения: поедая какое-либо крупное насекомое, ящерица неожиданно его оставляет и начинает методически схватывать и поедать находящихся в поле ее зрения мелких животных, например тлей, после чего снова принимается за оставленную крупную добычу. Подобное явление, называемое "сместительными реакциями", известно также у ряда других видов ящериц и у птиц. Интересно, что в некоторых исследованных нами желудках находились отдельные части довольно больших бабочек, жуков и дождевых червей, что, видимо, является следствием одновременного схватывания крупной добычи сразу несколькими особями. Подобные факты неоднократно удается наблюдать и в природе, причем нередко добыча оказывается настолько крупной, что насыщает 2-3 ящериц. Хотя широкая избирательность в отношении добычи у скальных ящериц отсутствует, на характере их питания в известной мере сказывается постепенно вырабатывающаяся привычка к поеданию определенного сорта добычи в период ее повышенной численности. Так, по окончании массового лета крылатых муравьев эти насекомые еще некоторое время продолжают занимать определенное место в пищевом рационе ящериц, "по инерции" предпочитающих их остальной добыче.
Имеющиеся в литературе сведения о пище ряда других форм скальных ящериц содержатся в табл. 34. Сравнение их с нашими данными указывает прежде всего на значительную однородность в питании отдельных видов и подвидов, причем наблюдающиеся количественные различия определяются, видимо, неодинаковым соотношением различных сортов добычи в разных биоценозах и в разное время года.
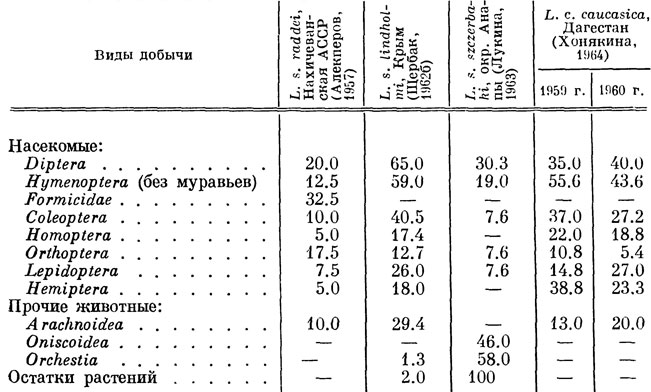
Таблица 34. Сравнительная встречаемость различных видов добычи в рационе некоторых форм скальных ящериц, %
Вместе с тем, помимо общих компонентов питания, для отдельных популяций ящериц характерны также присущие им свои виды добычи, определяемые спецификой местообитания или особенностями биологии данной формы. Например, L. s. nairensis, обитающие вблизи водоемов, в определенное время года в значительной мере переключаются на появляющиеся в массе имагинальные стадии развивающихся в воде насекомых, в частности ручейников и комаров-хирономусов. Хорошим примером такого рода служит обитающая на берегу моря, в непосредственной близости от воды, L. s. szczerbaki, пища которой, по данным Г. П. Лукиной (1963), в значительной мере слагается из живущих в зоне прибоя бокоплавов рода Orchestia. Аналогичным образом в пище высокогорной L. s. valentini большое место занимают многочисленные здесь саранчовые и уховертки.
Интересно, что во всех исследованных Лукиной (1963) желудках L. s. szczerbaki были обнаружены трудно определимые растительные остатки, что едва ли можно объяснить случайным заглатыванием их вместе с насекомыми. Отметим, что нормальное поедание растений обнаружено Радовановичем (Radovanovic, 1959) и у встречающейся на морских побережьях европейской стенной ящерицы L. melisellensis.
По имеющимся данным, интенсивность питания скальных ящериц, определяемая по количеству съедаемой за сутки добычи, заметно меняется по сезонам. Как показала З. П. Хонякина (1964), изучившая на большом материале питание L. caucasica в Дагестане, максимальное количество пустых желудков у половозрелых особей приходится на апрель (29.75%), май (33.3%), сентябрь (52.0%) и октябрь (42.8%), тогда как наименьшее наблюдается в июле (8.6%) и августе (14.2%). Таким образом, у исследованного вида максимальная интенсивность питания имеет место сразу после размножения, что биологически вполне оправдано, если учесть необходимость быстрого восполнения энергетических затрат, связанных с репродуктивной функцией.
По нашим данным, среднее за июль-август наполнение желудков у половозрелых особей L. armeniaca и L. s. valentini составляло соответственно 10.9% к весу тела для первых и 12.3 % для вторых при максимальном весе съеденной за сутки добычи 600-850 мг.
Наблюдающееся у половозрелых самок низкое в сравнении с самцами наполнение желудков объясняется резко пониженной интенсивностью питания в период размножения. Так, у большинства исследованных особей с яйцами в яйцеводах желудки были совершенно пустыми.
О повышенной интенсивности питания скальных ящериц в конце лета косвенно можно судить по ходу сезонной динамики развития жировых тел, вес которых резко возрастает сразу по окончании периода размножения, что в значительной мере связано также с прекращением функции гонад.
Энергетическая сторона питания скальных ящериц практически не изучена. В данной связи представляют интерес полученные нами при вольерном содержании некоторые данные о скорости переваривания и прохождения пищи через желудочный тракт. В частности, было установлено, что съедаемые отдельными особями в 9-10 часов утра мучные черви и кузнечики полностью перевариваются в течение суток и к утру следующего дня желудок оказывается пустым. Используя некоторые приведенные выше данные учетов численности, можно, например, подсчитать, что обитающие на площади 1 га 15,83 особи L. s. saxicola съедают за сутки биомассу, равную примерно 12 800 мг, принимая средний вес поедаемой за сутки добычи 800 мг.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'