Клинальная изменчивость и вопрос о выделении подвидов
Представление о характере географической изменчивости большинства рассматриваемых здесь видовых и подвидовых форм скальных ящериц дают сводные таблицы, составленные на основании анализов отдельных выборок, происходящих из различных, более или менее разобщенных точек их ареалов. Биометрическая обработка этих данных позволяет судить о достоверности различий между отдельными признаками, так же как между сравниваемыми популяциями в целом. Наглядное представление о степени различия исследованных выборок дают сводные профили, отражающие отклонения каждого признака, выраженные в долях среднего квадратичного отклонения от найденного по взвешенному методу среднего арифметического значения данного признака (MΣ).
Рассмотрение таблиц и построенных на их основании профилей показало, что отдельные признаки не подвержены закономерной географической изменчивости вовсе, тогда как у ряда других изменения эти вполне отчетливы и носят клинальный характер. В частности у многих изученных форм выяснилось существование клинальной изменчивости числа чешуй вокруг середины тела (признак 4) и по средней линии горла (признак 5), т. е. именно тех признаков, которые широко используются специалистами при таксономических исследованиях ящериц сем. Lacertidae. Более или менее выраженные, нередко хаотические гетероклина л ьные изменения прослеживаются также и у ряда других меристических показателей чешуйчатого покрова. У одних форм, например у L. s. saxicola, нарастание числа чешуй вокруг середины тела происходит с востока на запад (рис. 11), у других, как у L. г. rudis, - с запада на восток (рис. 43), у третьих, L. s. brauneri, - с северо-востока на юго-запад (рис. 13). Широко распространен случай, когда направление изменчивости одних и тех же признаков различно в разных частях ареала или прослеживается лишь на отдельных его участках. Специальный интерес представляет случай круговой изменчивости, наблюдаемый на участке ареала у L. s. valentini (рис. 29). Как и следовало ожидать, четкая клинальная изменчивость обнаруживается обычно лишь на участках непрерывного ареала и отсутствует в случаях, когда ареал более или менее значительно разорван. В данной связи представляет интерес рассмотрение влияния изоляции на характер изменчивости отдельных форм. Выше, характеризуя распространение скальных ящериц в пределах Кавказа, мы уже говорили, что многие из них обладают хорошо выраженными разорванными ареалами, особенно характерными для всех партеногенетических видов. Ранее (Даревский, 1960а, 1966а) нами было показано, что в большинстве случаев такие разрывы можно увязать с изменениями климата ледниковых эпох или же с интенсивной вулканической деятельностью, широко развернувшейся на Кавказе в позднечетвертичное время. Таким образом, согласно данным палеогеографии, можно полагать, что период существования разорванных ареалов, а следовательно, и период изоляции обособленных популяций скальных ящериц насчитывает не более 10-12 тыс. лет. Один из наиболее четких разрывов такого рода имеет место в распростренении номинального подвида L. rudis, обширная изолированная популяция которого существует на северных склонах Большого Кавказского хребта в Чечено-Ингушетии. Как видно на рис. 43, исследованная выборка из этой популяции по ряду показателей фолидоза заметно отличается от выборок из Грузии, причем эти отличия, не выходящие, однако, за пределы изменчивости данного подвида, выпадают из ряда постепенных изменений, обусловленных клинальной изменчивостью. В частности, обращает на себя внимание значительная редукция числа зернышек между верхнересничными и надглазничными щитками (признак 7), ряд которых в 90% случаев бывает прерван. Столь же четкие, статистически достоверные отличия по ряду признаков наблюдаются в выборке из изолированной популяции L. c. caucasica, обитающей на боковом Андийском хребте в Чечено-Ингушетии, причем, как и в случае с L. rudis, здесь имеет место почти стопроцентная редукция ресничных зернышек (рис. 48). Приведенные примеры показывают, что географическая изоляция отдельных популяций скальных ящериц влечет за собой весьма значительные изменения некоторых морфологических признаков фолидоза. В связи с этим естественно возникает вопрос, не является ли высокий полиморфизм рассматриваемой группы прямым следствием дивергенции морфологических признаков под влиянием более или менее длительной географической изоляции. Вопрос этот тем более уместен, что, согласно взглядам, развиваемым рядом современных эволюционистов, именно пространственная изоляция является ведущим фактором расо- и видообразования (Майр, 1947; Мауг, 1963). Рассмотрение этого вопроса в свете имеющихся в нашем распоряжении данных показывает, что некоторые формы ящериц подрода Archaeolacerta действительно сложились под влиянием географической изоляции, что наблюдается, однако, лишь в случаях, когда достаточно обширная отделившаяся популяция эволюционировала в новых для себя специфических условиях среды. Именно таким путем, как мы увидим дальше, образовалась, по всей видимости, форма Lacerta caucasica, сложившаяся в условиях сравнительно сурового горного климата после отделения от обитающего в предгорьях теплолюбивого подвида L. s. daghestanica. Характерно также, что изолированные популяции ряда других форм (L. s. portschinskii, L. s. brauneri, L. s. raddei, L. r. rudis и др.), обитающие на разных высотах, морфологически более четко отличаются друг от друга, нежели в случаях, когда такая изоляция не сопровождается заметным разрывом по вертикали. Таким образом, придавая большое значение географической изоляции, мы вслед за рядом отечественных исследователей (Оленов, 1959; Шварц, 1966а, и др.) не считаем ее однако, решающим фактором формообразования, возникающего прежде всего как следствие реакции изолированной популяции на специфические условия среды.
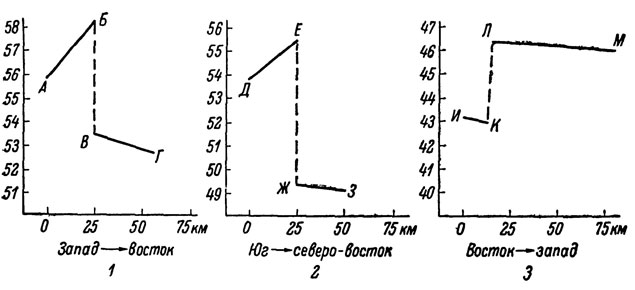
Рис. 65. Нарушение клинальной изменчивости среднего количества чешуй вокруг середины тела на стыках ареалов соседних подвидовых форм скальных ящериц: 1 - L. s. portschinskii - L. s. raddei; 2 - L. s. nairensis - L. s. valentini; 3 - L. c. cauvasica - L. c. alpina. A - Степанаван; Б, В - Туманян; Г - Иджеван; Д - Фантан; Е, Ж - Лчашен; З - Камо; И - ущелье Чегема; К. Л - Терскол; М - Теберда. По оси абсцисс - расстояние между географическими пунктами, отмеченными буквами
Как известно, термин "клина" и понятие клинальной изменчивости впервые были введены в литературу Гекели (Huxley, 1939) для обозначения широко наблюдающегося у животных постепенного географического изменения отдельных признаков. В последние годы явление клинальной изменчивости вновь привлекло к себе внимание некоторых таксономистов, склонных рассматривать самый факт ее существования как доказательство искусственности выделения географических подвидов. В отечественной литературе такая точка зрения последовательно развивается прежде всего П. В. Терентьевым (1957), который аргументировал ее также в ряде более поздних исследований (1966). Не рассматривая специально точку зрения П. В. Терентьева, заметим только, что, она, видимо, действительно справедлива в отношении ряда обладающих широкими ареалами видов, подвиды у которых нередко выделяются искусственно. Что же касается ящериц рассматриваемой группы, то ниже мы постараемся показать, что наличие хорошо выраженной клинальной изменчивости не только не препятствует, но, напротив, подчеркивает необходимость выделения у них объективных подвидовых форм. Уже Гекели (Huxley, 1939) отмечал, что характер клинальной изменчивости у разных видов бывает весьма различен, причем наряду с достаточно плавными клинами встречаются и крутые, характеризующиеся довольно резкими изменениями величин. Разнотипность клин, наблюдающихся на сплошных участках ареала, была показана позднее также многими другими исследователями. Например, в ряде работ, посвященных географической изменчивости oбурых лягушек, Л. Я. Топоркова (1964, 1966а, 19666) установила, что на фоне однотипных клинальных изменений могут быть выделены отдельные группы популяций, обладающие существенными морфологическими особенностями, позволяющими рассматривать их в качестве самостоятельных внутривидовых единиц. Выяснилось также, что области распространения таких форм приурочены к территориям со специфическими физико-географическими условиями, причем их морфологические отличия сопровождаются важными отличиями в экологии. Такая взаимосвязь вполне оправдана биологически, поскольку клины возникают в результате деятельности отбора и в большинстве случаев параллельны определяющему их градиенту факторов среды. Четкие нарушения постепенности клинальной изменчивости отдельных признаков, совпадающие с границами подвидов, наблюдаются также и у изученных нами видов скальных ящериц. Например, в северной Армении в зоне перехода между L. s. portschinskii и L. s. raddei имеет место отрицательный скачок количества чешуй вокруг середины тела, резко нарушающий плавный характер изменчивости этого признака от популяции к популяции у обоих перекрывающихся подвидов. Подобные же скачки обнаруживают этот и некоторые другие показатели, включая окраску, у ряда других подвидовых форм (рис. 65). Так, пограничные подвиды L. s. nairensis и L. s. valentini резко различаются, в частности, особенностями окраски: всегда зеленой спиной и желтым брюхом у первых и бурым или буровато-серым верхом и зеленым брюхом у вторых. В ряде случаев различия по отдельным признакам между подвидами в пограничной зоне выражены даже более четко, чем на периферии, в связи с чем их сравнительные диагнозы заметно различны для особей из разных частей ареалов.
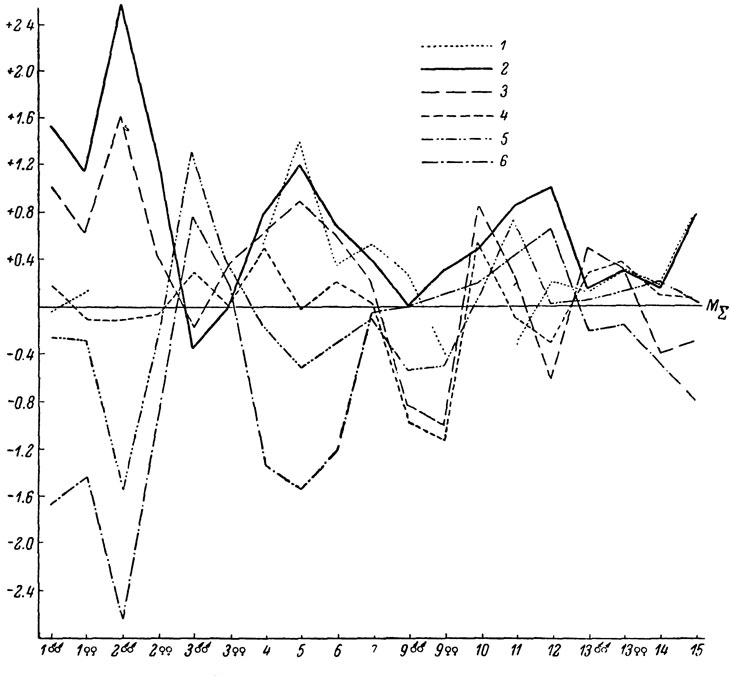
Рис. 66. Сводный профиль изменчивости Lacerta saxicola (северная группа подвидов): 1 - L. s. lindholmi; 2 - L. s. szczerbaki; 3 - L. s. darevshii; 4 - L. s. brauneri; 5 - L. s. saxicola; 6 - L. s. daghestanica
Особенно важно подчеркнуть, что такого рода резкие сдвиги или ступеньки (stepcline английских авторов) клинальной изменчивости часто совпадают с изменениями условий среды, в частности с характером местообитаний или границами вертикальных ландшафтных зон. Это можно видеть, например, у обитающих на Северном Кавказе L. s. saxicola и L. s. darevskii. Первая из этих ящериц встречается в междуречье Чегема и Теберды в пределах Кабардино-Балкарии и юга Ставрополья, тогда как вторая широко распространена главным образом в предгорьях Краснодарского края. Граница между обоими подвидами хорошо отвечает наблюдающейся здесь смене сухих предгорных ландшафтов Ставрополья лесными массивами Теберды и Краснодарского края (рис. 10). Во многих других случаях границы между подвидами определяются сменой зональности ландшафтов, как это наблюдается, в частности, в случае с предгорной формой L. s. nairensis и высокогорной L. s. valentini. Вследствие этого мы с полным правом можем говорить о предгорных, лесных, горностепных и альпийских подвидах скальных ящериц, существенно различающихся не только по своей морфологии, но также и по экологическим особенностям. Отметим, что аналогичная зависимость морфологических особенностей внутривидовых популяций от условий среды была установлена у полевок рода Clethrionomys (Большаков и Шварц, 1962) и показана, кроме того, у ряда других животных. Таким образом, нашим материалом полностью подкрепляется все более утверждающая себя в последнее время точка зрения, что в основу выделения подвидовых форм должна быть положена прежде всего специфическая эволюционная судьба части вида, определяющаяся спецификой среды ее обитания (Шварц, 1966а).
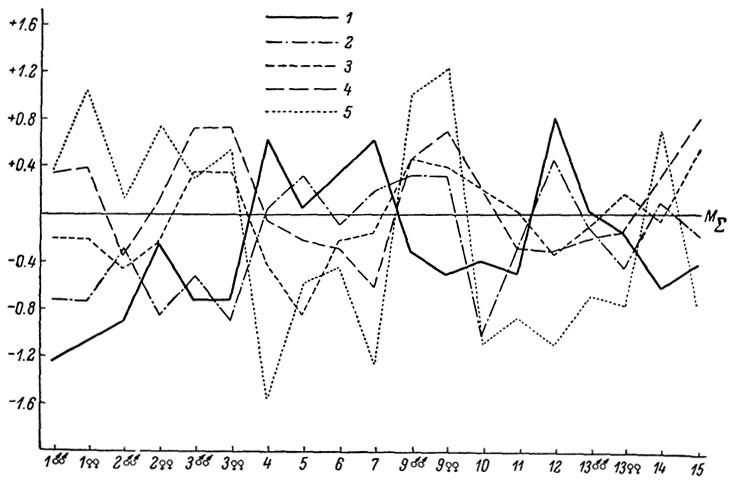
Рис. 67. Сводный профиль изменчивости L. saxicola (южная группа подвидов): 1 - L. s. parvula; 2 - L. s. portschinskii; 3 - L. s. raddei; 4 - L. s. nairensis; 5 - L. s. valentini
Интересно, что в отдельных случаях граница между соседними подвидами скальных ящериц определяется, видимо, какими-то еще не вполне ясными историческими причинами. Такова, например, проходящая на центральном Кавказе зона разграничения между высокогорными подвидами L. с. caucasica и L. с. alpina, первый из которых распространен строго к востоку, а второй - к западу от Эльбруса (рис. 48). Отметим, что в этом же районе наблюдается разделение ареалов и некоторых других высокогорных животных, например восточнокавказского (Capra cylindricornis) и западнокавказского (Capra caucasica) козлов (Верещагин, 1959).
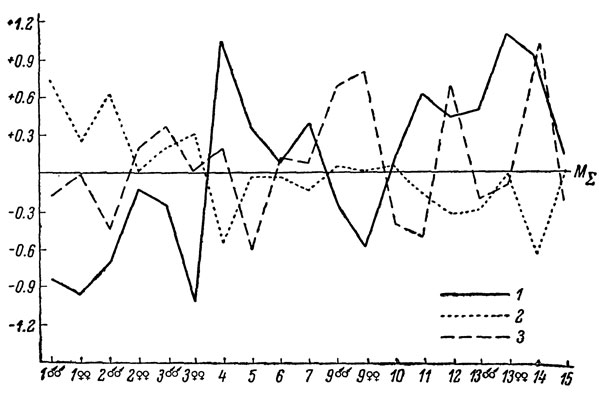
Рис. 68. Сводный профиль изменчивости Lacerta rudis: 1 - L. r. obscura; 2 - L. r. rudis; 3 - L. r. macromaculata
В целом характер внутривидовой изменчивости рассматриваемых нами полиморфных видов виден на сводных видовых профилях (рис. 66-69).
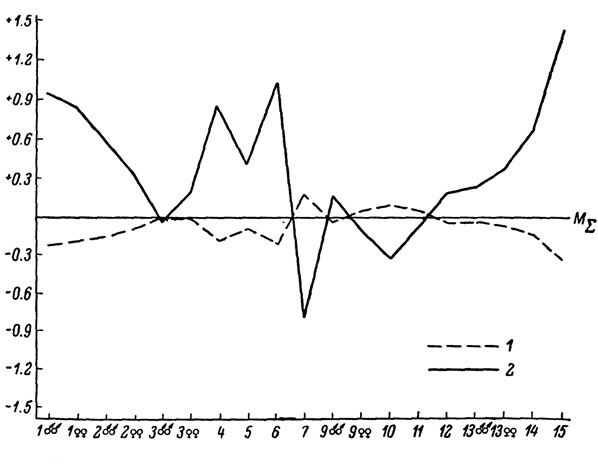
Рис. 69. Сводный профиль изменчивости Lacerta caucasica. 1 - L. c. caucasica; 2 - L. c. alpina
Анализ географической изменчивости всей группы кавказских скальных ящериц показывает, что клинальная изменчивость между видами по большинству признаков отсутствует. О степени различия всех исследованных видов можно судить по сводному профилю (рис. 70), из которого видно, что наиболее значительный размах изменчивости ряда меристических показателей фолидоза обнаруживается у сравнительно молодых форм L. caucasica, L. rudiso, L. armeniaca, L. rostombekovi, L. unisexualis и L. mixta. Что же касается исходного вида L. saxicola, то характеризующие его показатели, как и можно было ожидать, лишь незначительно отклоняются от среднего их значения. К этому необходимо добавить, что виды хорошо различаются также некоторыми особенностями расположения отдельных элементов чешуйчатого покрова, не нашедших своего отражения на сводном профиле. Так, L. unisexualis характеризуется статистически достоверным отсутствием шва между межчелюстным и лобоносовым щитками, L. armeniaca - характерным удвоением промежуточных щитков височной области, L. dahli - исчезновением или укорочением шва между задненосовым и лобоносовым щитками и т. д. Интересно отметить, что популяции разных подвидовых форм скальных ящериц нередко характеризуются присутствием единичных особей другого подвида, что может наблюдаться в любой точке ареала независимо от наличия или отсутствия зоны интерградации. Так, в популяциях номинальной формы L. caucasica встречаются иногда отдельные особи L. c. alpina1 а в популяциях L. s. darevskii - особи L. s. brauneri. Отметим, что, по С. С. Шварцу (1959, 1966а), такое явление довольно часто наблюдается у подвидов многих животных и его следует рассматривать как одну из форм обратимости внутривидовой изменчивости.
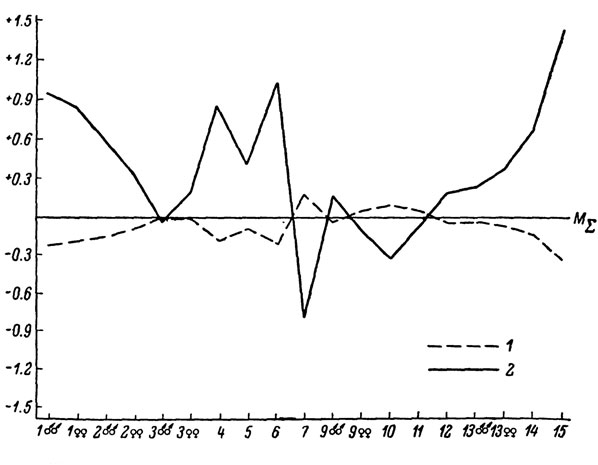
Рис. 70. Сводный профиль изменчивости кавказских видов скальных ящериц подрода Atchaeolacerta: 1 - L. armeniaca; 2 - L. dahli; 3 - L. rostombekovi; 4 - L. unisexualis; 5 - L. saxicola; 6 - L. rudis; 7 - L. caucasica; 8 - L. mixta
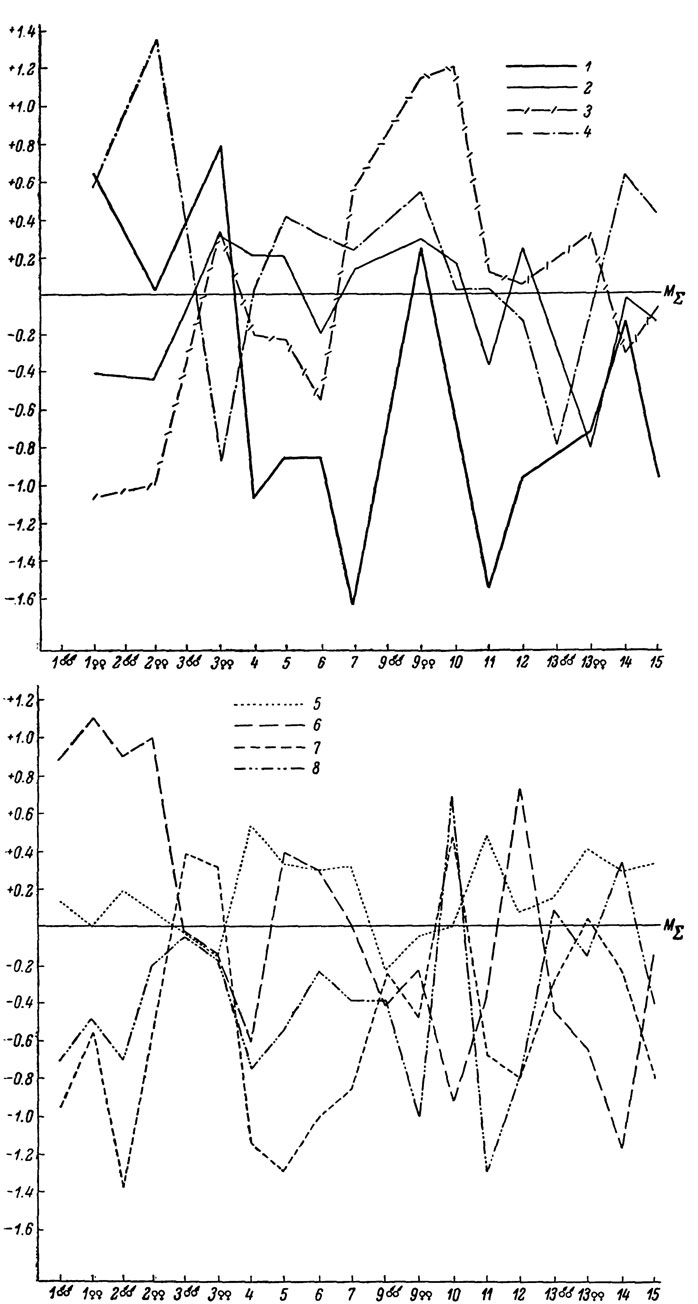
Рис. 70. Сводный профиль изменчивости кавказских видов скальных ящериц подрода Atchaeolacerta: 1 - L. armeniaca; 2 - L. dahli; 3 - L. rostombekovi; 4 - L. unisexualis; 5 - L. saxicola; 6 - L. rudis; 7 - L. caucasica; 8 - L. mixta
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'