Специфика изменчивости партеногенетических видов
Как и следовало ожидать, благодаря односторонней материнской наследственности фенотипическая изменчивость у всех партеногенетических видов скальных ящериц оказалась гораздо меньшей, чем у близких обоеполых форм. Это хорошо видно из табл. 28, где статистически вычислен коэффициент вариации С% для двух важных в систематическом отношении признаков - количества чешуек вокруг середины тела Sq и количества чешуек по средней линии горла G. Можно видеть, что у партеногенетических форм коэффициент вариации значительно ниже, чем у бисексуальных, что в равной мере относится и к ряду других признаков, включая окраску тела, благодаря чему партеногенетические сестры из каждого выводка (клона) более похожи друг на друга и на свою мать, чем это имеет место у обоеполых видов. Кроме того, благодаря малой фенотипической изменчивости внутри каждого отдельного клона пределы варьирования одних и тех же признаков внутри партеногенетических популяций в целом также сравнительно невелики, несмотря на множество мелких фенотипических линий. В связи с этим партеногенетические виды сравнительно мало изменчивы, что можно видеть при сравнении средних квадратных отклонений отдельных признаков в сводных таблицах, а также сводных профилей, характеризующих изменчивость всех рассматриваемых здесь обоеполых и партеногенетических форм. Отметим, что более узкая фенотипическая изменчивость североамериканских партеногенетических ящериц Cnemidophorus velox в сравнении с близким бисексуальным видом C. inornatus была показана в последнее время Тэйлором (Taylor, 1965). Аналогичные данные для изолированных популяций партеногенетического вида С. tessellatus приводит Цвейфель (Zweifel, 1965).
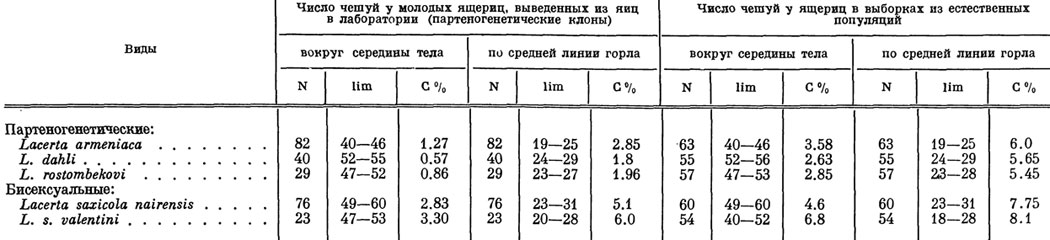
Таблица 28. Некоторые различия фенотипической изменчивости партеногенетических и бисексуальных форм скальных ящериц (по Даревскому и Куликовой, 1961)
Заслуживает внимания также тот факт, что благодаря большей морфологической однородности молодняка внутри каждого отдельного выводка фенотипическая изменчивость ящериц, выведенных из яиц в лаборатории, в целом оказалось заметно ниже, чем у особей из выборок, взятых в природе, что в равной мере относится к партеногенетическим и бисексуальным подвидам. Это показывает, что слагающиеся из многочисленных клонов естественные "популяции" партеногенетических ящериц в целом обладают достаточно высокой изменчивостью, способствующей их повышенной стойкости к изменениям во внешней среде. Обращают на себя внимание также некоторые свойственные партеногенетическим формам специфические особенности окраски, относящиеся прежде всего к расцветке нижней стороны тела. Если в окраске брюха большинства бисексуальных форм преобладают яркие желтые, оранжевые, зеленые или кирпично-красные тона, то у партеногенетических ящериц брюхо желтоватое или даже совершенно белое. Видимо, это объясняется исчезновением роли полового диморфизма в окраске как сигнального фактора, имеющего значение для обнаружения полами друг друга при спаривании.
Значительный интерес представляют собой также характеризующие партеногенетических ящериц особенности чешуйчатого покрова, послужившие в свое время основанием к описанию их в качестве особых подвидов. Так, у L. armeniaca имеет место своеобразное парное расположение некоторых щитков височной области (рис. 52, В, Д), у L. dahli наблюдается появление второго задненосового щитка или значительное укорочение шва между лобоносовым и задненосовым щитками (рис. 55, Б), у L. rostombekovi происходит значительное уменьшение преанальных щитков (рис. 57, Е), а у L. unisexualis появляется отсутствующий в норме широкий шов, соединяющий межчелюстной и пред лобный щитки (рис. 59, А). Характерно, что все эти особенности, наблюдающиеся у 99.8% партено-генетических особей, мало характерны для бисексуальных подвидов скальной ящерицы или во всяком случае не служат для них диагностическими признаками. С другой стороны, такие же диагностические признаки свойственны ряду филогенетически более древних южноевропейских ящериц подрода Archaeolacerta. Например, характерное для L. dahli укорочение или исчезновение шва между носовым и лобоносовым щитками или появление второго задненосового в норме свойственно балканской L. horvathi Mehely, для которой присуще также обычное наличие шва между предлобным и межчелюстными щитками, как это имеет место и у партеногенетической L. unisexualis. Как мы увидим дальше, такое своеобразное проявление у партеногенетических ящериц ряда атавистических признаков представляет определенный интерес при выяснении филогенетических связей внутри всей группы стенных-скальных ящериц в целом.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'