Изменчивость в зонах совмещения ареалов
Мы уже говорили, что специфической особенностью распространения скальных ящериц в пределах Кавказа, и, видимо, также и Малой Азии является наблюдающееся у них частичное перекрывание ареалов. При этом нужно различать случаи, когда друг на друга в той или иной мере налегают области распространения соседних подвидов или же совмещенными на значительном протяжении оказываются ареалы соседних видов. Как показывают наши исследования, в первом из этих случаев образующиеся зоны совместного обитания чаще всего связаны со спецификой вертикального распространения ящериц и обнаруживаются обычно в ущельях горных рек, по которым формы, распространенные в предгорьях, как бы вклиниваются в область распространения форм, ареал которых располагается выше. При этом численность "вклинивающейся" формы в зоне перекрывания бывает, как правило, ниже, чем в ее основном ареале. В случаях же совмещения ареалов широко распространенных видов, например L. caucasica и L. rudis, численность каждого из них в смешанных популяциях примерно одинакова. Известны также сравнительно небольшие смешанные популяции, слагающиеся из трех разных видов и подвидов, причем в этом случае обычно наблюдается лишь незначительная примесь третьей формы, уступающей в численности двум основным. Подобные случаи известны, например, в ущелье р. Дзорагета в северной Армении, где "вклинивающаяся" L. s. portschinskii встречается с L. armeniaca и L. dahli, и на северо-западных склонах Триалетского хребта в окр. Бакуриани в Грузии, где вместе с L. mixta и L. armeniaca встречаются отдельные особи L. r. obscura.
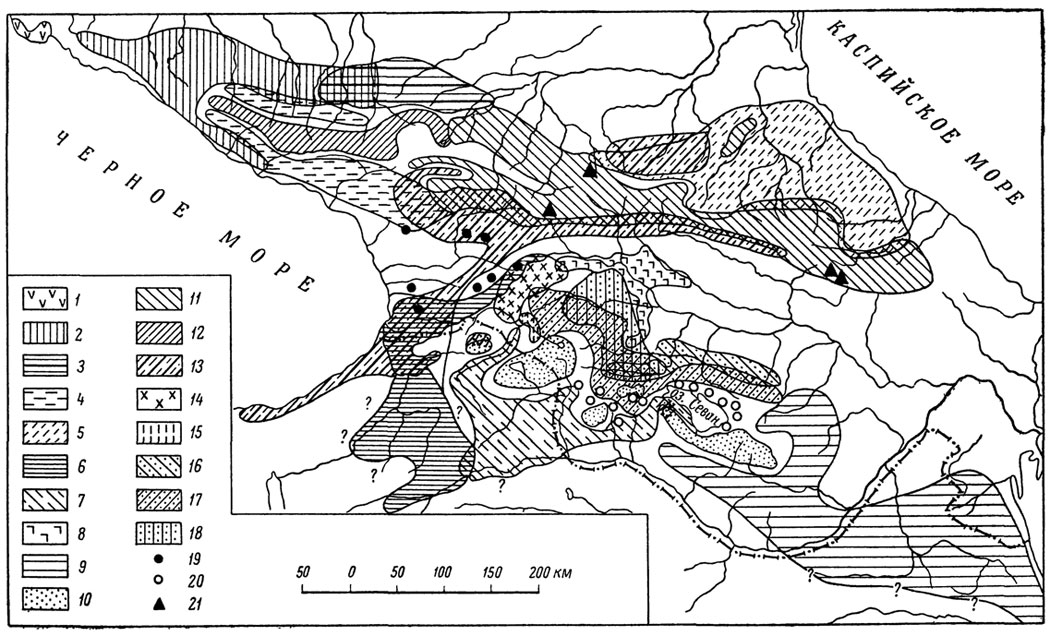
Рис. 64. Сводная карта основных ареалов скальных ящериц в пределах Кавказа: 1 - L. saxicola szczerbaki, 2 - L. s. darevskii, 3 - L. s. saxicola; 4 - L. s. brauneri; 5 - L. s. daghestanica; 6 - L. s. parvula; 7 - L. s. nairensis; 8 - L. s. porischinshii; 9 - L. s. raddei; 10 - L. s. valentini; 11 - L. caucasica caucasica; 12 - L. c. alpina; 13 - L. rudisrudis; 14 - L. r. obscura; 15 - L, r. macroinaculata; 16 - L. rostombekovi; 17 - L. armeninca; 18 - L. dahli; 19 - основные местонахождения L. mixta; 20 - основные местонахождения L. unisexualis; 21 - изолированное местонахождение L. s. daghestanica
Важный вопрос о механизмах репродуктивной изоляции, действующих в смешанных популяциях скальных ящериц, будет специально рассмотрен ниже. Можно отметить, что аналогичные зоны совмещения ареалов различных видовых и подвидовых форм из группы европейских стенных ящериц описаны из ряда мест Южной Европы и западной части Малой Азии (Mertens, 1952; Klemmer, 1957).
Что касается скальных ящериц, то основные, известные в настоящее время в пределах Кавказа зоны совмещения ареалов приведены в табл. 27.

Таблица 27. Основные зоны частичного совмещения ареалов скальных ящериц в пределах Кавказа
Характерное для большинства кавказских форм перекрывание подвидовых и видовых ареалов отражено также на сводной карте рис. 64. Как показывают наши исследования, наблюдающиеся зоны совмещения ареалов соседних подвидовых форм характеризуются обычным присутствием в них так называемых промежуточных особей, которых на основании морфологического критерия не удается достоверно отнести ни к первому, ни ко второму подвиду. Такое явление имеет место, в частности, на Северном Кавказе в зонах совмещения ареалов L. s. darevskii и L. s. brauneri и L. s. darevskii и L. s. saxicola (рис. 1). Можно отметить, что такого же рода более или менее выраженные "классические" переходы между подвидами известны и у многих других животных, причем существование в смешанных популяциях особей, сочетающих в себе признаки обоих симпатрических видов, в равной мере можно объяснить как первичной, так и вторичной интерградацией. Не останавливаясь на многочисленных примерах такого рода, приводимых, в частности, Майром (1947), сошлемся лишь на чрезвычайно интересную работу Савада (Sawada, 1963), показавшего существование четких промежуточных популяций у японского тритона (Triturus pyrrogaster). Теоретические аспекты этого вопроса в общей форме были рассмотрены недавно также Н. И. Лариной и В. П. Денисовым (1966). Особый интерес представляет широко распространенный случай, когда на значительном протяжении оказываются совмещенными ареалы двух хорошо различающихся видов, например L. saxicola и L. rudis или L. rudis и L. caucasica (рис. 2). Как и следовало ожидать, какие бы то ни было промежуточные формы между такими полусимпатрическими видами отсутствуют, что, как мы увидим дальше, объясняется действием биологических изолирующих механизмов, заключающихся в разновременности созревания гонад и различии сигнальной брачной окраски.
Предпринятое нами специальное изучение этого вопроса показало однако, что в ряде случаев ящерицы из смешанных популяций морфологически заметно отличаются от особей своего же вида из чистых аллопатрических участков его ареала. Такие различия можно видеть на примере L. s. parvula, значительная часть ареала которой перекрывается с областью распространения L. rudis. Как видно из табл. 12 и рис. 38, выборка из чистой популяции этого подвида в северо-восточной Турции по ряду признаков хорошо отличается от выборок из смешанных популяций из Аджарии и соседних районов Грузии, причем различия эти заключаются в заметном отрицательном отклонении всех меристических показателей фолидоза. Иными словами, L. s. parvula, обитающая совместно с L. rudis, характеризуется более крупным масштабом элементов чешуйчатого покроваг не выходящим, однако, за пределы изменчивости, установленной для данного подвида. Подобного рода более или менее выраженные отклонения меристических признаков наблюдаются и в смешанных популяциях некоторых других форм, чем в значительной мере объясняется отмеченное выше смазывание у них общей картины клинальной изменчивости. По нашему мнению, это происходит вследствие некоторого общего изменения фенотипа гетерогенных популяций под воздействием единичных случаев перекрестного обмена гамет, происходящих в обход механизмов репродуктивной изоляции, о которых уже говорилось выше. Такие случаи могут, например, иметь место при нередко наблюдающихся индивидуальных сдвигах в сроках размножения отдельных особой. Возникающие при этом гибридные генотипы быстро растворяются в исходном генотипе родительских форм, однако накопление их за длительное время сопряженного существования приводит постепенно к заметному общему изменению фенотипа смешанных популяций. Возникающие таким образом фенотипические отклонения могут вырастать до ранга едва ли не достоверных таксономических различий, как это наблюдается, например, у L. s. parvula. Поскольку в этом случае обладающие совмещенными ареалами L. rudis и L. s. parvula по величине значительно отличаются друг от другаг перекрестное их оплодотворение происходит лишь в одном направлении,, а именно путем спаривания молодых половозрелых самцов L. rudis со взрослыми самками L. s. parvula. Именно поэтому накопившиеся изменения генотипа сказались лишь на морфологии материнской формы и почти не коснулись отцовской. Однако в подобной же ситуации при сопряженном обитании близких по величине L. s. saxicola и L. c. caucasica или L. caucasica и L. s. brauneri фенотипы каждой из них оказались несколько сдвинутыми в сравнении с несовмещенными частями их популяций (рис. 12 и 15). Отметим, что подобное же явление было описано в свое время С. Д. Перелешиным (1928) у прыткой ящерицы в зоне совмещения ареалов L. a. agilis и L. a. exigua. Установленое нами изменение фенотипа симпатриантов в смешанных популяциях скальных ящериц очень напоминает описанное Стугреном и Поповичем явление так называемой интерклинальной изменчивости, констатированное ими в зонах совмещения видовых ареалов желтобрюхой и краснобрюхой жерлянок (Стугрен и Попович, 1961; Stugren, 1962). Отметим, что, по данным этих авторов, существующие на территории Румынии гетерогенные популяции Bombina bombina и Bombina variegata, так же как в нашем случае с ящерицами, не образуют особого подвида, а четко детерминируются как первый или второй вид.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'