2. Орган обоняния предков наземных позвоночных и условия его формирования
Прежде всего необходимо установить, что было ведущим процессом в ходе эволюционных преобразований органа обоняния, позволившим водным животным использовать этот орган в совершенно иной, воздушно-наземной, среде. У всех современных, обладающих хоанами, позвоночных, за исключением хвостатых и бесхвостых амфибий, существует первоначальная связь между ноздрей и хоаной, т. е. вводным и выводным носовыми отверстиями, в виде оро-назальной борозды, соединяющей в эмбриональном состоянии обонятельную и ротовую ямки и превращающейся затем в хоанальный канал. Это обстоятельство наряду с наличием у костных рыб двух наружных ноздрей (передней - вводной и задней - выводной) уже давно привело к мысли о том, что хоана наземных позвоночных обязана своим происхождением перемещению в ротовую полость одной из наружных ноздрей, а именно передней (А. Н. Северцов, 1926а). Причины такого перемещения А. Н. Северцов не касался. В сравнительно недавнее время вопрос об отношении хоаны наземных позвоночных к наружным носовым отверстиям рыб был подробно разобран И. И. Шмальгаузеном (1957б, 1958д, 1960, 1964б), который обосновал и возможные причины смещения ноздрей в ротовую полость. По И. И. Шмальгаузену этот процесс не имеет никакого отношения к приспособлению для воздушного дыхания (подобные мысли высказывались и ранее: Карпилов, 1940; Atz, 1952b; Татаринов, 1953), а связан с образом жизни кистеперых предков наземных позвоночных, которые были малоподвижными донными подстерегающими хищниками. Положение ноздрей у края рта усиливало интенсивность просасывания воды через орган обоняния при дыхательных движениях рыбы и тем самым увеличивало эффективность его работы. Наибольшей скорости ток воды достиг после перемещения задней ноздри, точнее ее медиальной части, внутрь ротовой полости. Эта гипотеза подтверждается сохранившимися следами перемещения хоаны внутрь ротовой полости у бесхвостых амфибий от исходного, более наружного положения.
Что касается хвостатых амфибий, то следов перемещения хоаны у них не сохранилось вследствие вторично измененного развития ротовой полости, но зато наблюдается эмбриональное перемещение ноздрей из исходного положения на конце морды по ее вентральной поверхности к ротовому впячиванию, чем достигается очень близкое расположение ноздри и рта в момент образования хоанального канала, и последующий обратный сдвиг ноздрей вперед и дальше на дорсальную сторону головы (рис. 77). Причем эти перемещения более замедлены и легче прослеживаются как раз у самых примитивных хвостатых амфибий - Hynobius keyserlingii и в особенности Ranodon sibiricus, выступая в более стертой форме у высших Urodela, что указывает на подобное перемещение передних ноздрей у их предков. В связи с этим чрезвычайно интересно, что у примитивных стегоцефалов Ichthyostega и Acanthostega ноздри смещены к краю рта, вследствие чего инфраорбитальный сейсмосенсорный канал оттеснен внутрь (Jarvik, 1952, 1955). Ярвик считает такое положение ноздрей у Ichthyostegalia вторичным. Однако Панчен (Panchen, 1964, 1967), произведя ревизию ряда нижнекарбоновых лабиринтодонтов, в том числе Palaeogyrinus, Megalocephalus и других, приходит к выводу (Panchen, 1967), что в четырех примитивных группах (Ichthyostegidae, Acanthostegidae, Anthracosauria и Loxommidae) все достаточно хорошо сохранившиеся экземпляры имеют наружные ноздри, располагающиеся у края рта, и что это было примитивным состоянием для лабиринтодонтов и, возможно, для наземных позвоночных вообще. Тенденция к смещению ноздрей к краю рта наблюдается уже у некоторых кистеперых рыб, например у Panderichthys, который по ряду признаков наиболее близок к тетраподам (Воробьева, 1960а, 1960б, 1962, 1971а, 1971б; Schultze, 1969). У всех же более поздних форм стегоцефалов ноздри располагаются гораздо дальше от края рта, на дорсальной поверхности головы.
Следовательно, происходящее в онтогенезе хвостатых амфибий передвижение ноздрей сначала к краю рта, а затем на дорсальную сторону морды находит свое подтверждение в палеонтологических данных и делает гипотезу И. И. Шмальгаузена (1957б) о подобном филогенетическом перемещении обеих ноздрей (как передней, так и задней) кистеперых рыб к краю челюсти и последующем возвращении передней ноздри на дорсальную сторону переднего конца головы очень достоверной. Упрек Ярвика (Jarvik, 1964) в адрес И. И. Шмальгаузена, что его гипотеза ошибочна, потому что он пользовался устаревшими реконструкциями Osteolepis и Ichthyostega, на которых инфраорбитальный сейсмосенсорный канал изображался доходящим только до ноздри, тогда как по новейшим данным этот канал проходит у них без перерыва под ноздрей, верен только в отношении, касающемся несовершенства реконструкций. Но и в этом случае факт положения ноздрей на краю рта у примитивных стегоцефалов и уже у некоторых кистеперых рыб остается незыблемым. Кроме того, И. И. Шмальгаузен (1955в, 1957б, 1958д) построил свою гипотезу не на палеонтологических реконструкциях, которыми сам никогда не занимался, а на собственных эмбриологических исследованиях развития инфраорбитальной сейсмосен-сорной линии и иннервирующего эту линию щечного нерва (n. buccalis VII) у наиболее примитивных хвостатых амфибий - Hynobius keyserlingii и Ranodon sibiricus. Ход развития у Hynobiidae инфраорбитальной сейсмо-сенсорной плакоды показывает, что непрерывность этой линии, существующая у современных костных рыб (Amia, Lepidosteus), у примитивных хвостатых амфибий нарушена: премаксиллярная часть инфраорбитальной линии развивается у Hynobiidae из отдельной плакоды и снабжается особой ветвью щечного нерва, которая в отличие от общего поверхностного прохождения нервов сейсмосенсорных органов располагается глубоко, обходя с медиальной стороны обонятельный мешок; субназальная часть этой же линии также снабжается особыми, более длинными веточками щечного нерва (И. И. Шмальгаузен, 1957б, рис. 9 и 10). Такое развитие как самой инфраорбитальной линии, так и снабжающего ее нерва возможно только при вторичном восстановлении единства линии после произошедшего разрыва. Для современных хвостатых амфибий вообще характерно вторичное разрастание линий сейсмосенсорных органов: у аксолотля, например, та же субназальная часть инфраорбитальной линии, разрастаясь, вытесняет первичную премаксиллярную часть и вызывает тем самым редукцию относящейся к ней ветви щечного нерва.
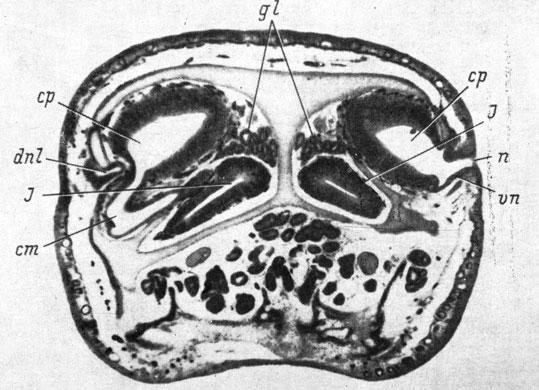
Рис. 9. Поперечный срез органа обоняния Rana temporaria (увел. 40)
Сопоставление всех этих данных позволило И. И. Шмальгаузену сделать вывод о нарушении целостности прохождения инфраорбитальной линии, обязанном своим происхождением тому, что эта линия была прервана задней ноздрей при перемещении последней в ротовую полость. Отсюда становится вполне понятным необычный ход щечного нерва у примитивных хвостатых амфибий, вызванный сопутствовавшим перемещению ноздри поворотом обонятельного мешка, сместившего нерв в глубину. Отдельная закладка премаксиллярной части инфраорбитальной линии, особая иннервация субназальной части, показывающая ее вторичное разрастание после разрыва, являются также чрезвычайно убедительными доказательствами именно этого пути возникновения хоаны наземных позвоночных. То, что привлеченные И. И. Шмальгаузеном для подкрепления своей гипотезы палеонтологические данные при последующей ревизии оказались несостоятельными, не опровергает этой гипотезы, получившей еще ряд подтверждений в настоящем исследовании, так как ни Osteolepis, ни Ichthyostega не являются прямыми предками наземных позвоночных. Отсутствие палеонтологического подтверждения вообще не может поставить под сомнение результаты эмбриологического исследования современных форм, поскольку первое зависит не от реальности фактов, а от случайности находок и правильности их интерпретации. К тому же И. И. Шмальгаузен (1957б) ссылается также и на другие формы примитивных стегоцефалов, у которых инфраорбитальный канал кончается у заднего края ноздри, например на Megalocephalus и Palaeogyrinus (Watson, 1926), и на двоякодышащих рыб, у которых и по новейшим исследованиям, подтверждающим более раннюю работу Хольмгрена и Пирсона (Holmgren а. Pehrson, 1949), ход инфраорбитального канала прерван ноздрями (Bertmar, 1966b), о чем в свое время писал и сам Ярвик (Jarvik, 1955). Однако в данном случае (Jarvik, 1964) он об этих фактах не вспоминает, хотя они пока не опровергнуты, а И. И. Шмальгаузен приводил их в сравнение для доказательства того, что инфраорбитальный канал может быть прерван при передвижении ноздрей.
Из всего сказанного следует, что хоана хвостатых амфибий, образовавшаяся из задней ноздри рыб, должна была иметь первоначальную связь с органом обоняния, так же как хоаны всех остальных Tetrapoda. С другой стороны, эмбриональное развитие органа обоняния хвостатых и бесхвостых амфибий показывает, что он образуется из двух морфологически обособленных закладок, одной из которых является сама обонятельная плакода, а другой - хоана с частью хоанального канала. Единственным логическим выводом из сопоставления всех этих данных может быть только следующий: в процессе исторического развития органа обоняния хвостатых и бесхвостых амфибий произошел разрыв эмбрионального органа обоняния на обонятельный мешок с частью хоанального канала, с одной стороны, и хоану с прилежащей к ней частью хоанального канала - с другой. Сразу же возникают вопросы: как и когда это могло произойти и почему это произошло только у хвостатых и бесхвостых амфибий?
Известно, что как для позднейших стегоцефалов, так и для современных хвостатых и бесхвостых амфибий характерно вторичное разрастание сейсмосенсорных линий. Такое разрастание именно инфраорбитальной линии и вторичное образование ее субназальной части доказано И. И. Шмальгаузеном (1955в, 1957б) уже для примитивных хвостатых амфибий, а у высших оно достигает еще большей степени. Но ведь в процессе эмбрионального развития закладка инфраорбитальной линии у Hynobiidae и других хвостатых амфибий, и именно субназальной части этой линии, вдвигается между органом обоняния и ротовым впячиванием еще до возникновения хоанального канала и тем самым препятствует его образованию на поверхности тела (рис. 19, А; 20, А; 23, Б). Это-то вторичное разрастание инфраорбитальной линии и послужило причиной разрыва закладки органа обоняния на две морфологически обособленные части, и произошло оно после филогенетического перемещения задней ноздри в ротовую полость. Этому имеются и экспериментальные доказательства. Презумптивный материал хоанального канала располагается именно в той части обонятельной плакоды, которая прилежит к инфраорбитальной плакоде органов боковой линии (рис. 41, А), отделяющей обонятельную плакоду от места ротового впячивания. Такое местоположение материала хоанального канала подтверждает, что причиной разрыва хоанального канала у предков хвостатых и бесхвостых амфибий было врастание инфраорбитальной плакоды между обонятельной плакодой и ротовым впячиванием. Результатом этого разрыва явилось то, что образующийся хоанальный канал был принужден разрастаться не на поверхности тела в виде оро-назальной борозды, а расти вглубь в виде выроста обонятельного мешка. Тем самым совершенно изменился и способ развития органа обоняния. У современных хвостатых и бесхвостых амфибий нет оро-назальной борозды, но она была у их предков, так же как она имеется и сейчас у всех остальных современных позвоночных, обладающих хоанами.
Что же могло быть причиной того, что это разрастание инфраорбитальной линии, вызвавшее разрыв закладки органа обоняния на две отдельные части, произошло только у предков современных хвостатых и бесхвостых амфибий и не затронуло остальных Tetrapoda, в том числе и гимнофион? И. И. Шмальгаузен (1955б, 1957а, 1958а, 1960) анализом сравнительно-анатомических данных, имеющихся по ископаемым кистеперым рыбам и стегоцефалам, показал, что большинство стегоцефалов переходило ко вторичноводному образу жизни. Черты организации современных хвостатых и бесхвостых амфибий также носят на себе следы вторичного возврата в воду. Одним из таких следов и является вторичное разрастание сейсмосенсорных линий, часто сопровождающееся и вторичным размножением чувствующих почек, которое наблюдается даже у самых примитивных современных хвостатых амфибий - Hynobiidae. Наряду со вторичным возвратом в воду, послужившим причиной разрастания линий сейсмосенсорных органов у взрослых животных, для хвостатых и бесхвостых амфибий вообще характерно продление личиночного развития в воде, во многих случаях приведшее у этих групп к возникновению целого ряда неотенических форм (Ambystomidae, Proteidae и др.). Не являлись редкостью неотенические формы и у лабиринтодонтов, например, Dvinosaurus (Быстров, 1938), Tungussogyrinus (Ефремов, 1939), целая группа Phyllospondyli, которых Ромер (Romer, 1939, 1964) хотя и считает только личинками различных рахитомов, но большинство исследователей выделяет в отдельную самостоятельную группу (Heyler, 1954, 1957; Lehman, 1955; Конжукова, 1964).
Продление личиночной жизни, протекающей в воде, и увеличение в результате этого биологического значения органов боковой линии у личинки должно было вызвать вторичное разрастание сейсмосенсорных линий уже в раннем развитии хвостатых и бесхвостых амфибий. У неотенических форм оно достигает максимума - примером может служить аксолотль, у которого вторичная субназальная часть инфраорбитальной линии, разрастаясь, даже вытесняет первичную премаксиллярную часть, что в свою очередь влечет за собой редукцию соответствующей ветви щечного нерва (И. И. Шмальгаузен, 1957б). С другой стороны, предки как гимнофион, так и остальных современных Tetrapoda, осваивая сушу, вообще утрачивали личиночное развитие в воде и переходили к иному способу размножения путем откладки яиц, защищенных прочными оболочками. Развитие внутри таких яиц несомненно способствовало редукции сейсмосенсорных линий, а не их вторичному разрастанию и тем самым обеспечило сохранение оро-назальной борозды, а следовательно, и морфологического единства эмбрионального органа обоняния. Общеизвестно, что у высших наземных позвоночных сейсмосенсорная система вообще редуцируется. Что же касается гимнофион, то у них сейсмосенсорные линии закладываются позже, чем у хвостатых и бесхвостых амфибий, в то время когда хоаны уже образовались (P. Sarasin u. F. Sarasin, 1887, 1890; Brauer, 1899). Во всяком случае образование у современных Tetrapoda, за исключением хвостатых и бесхвостых амфибий, в процессе развития органа обоняния оро-назальной борозды служит лучшим доказательством того, что разрастания инфраорбитальной линии до возникновения хоанального канала у них не было.
Оказавшаяся в результате всех этих сложных перемещений в ротовой полости задняя ноздря кистеперых рыб обеспечила более интенсивное функционирование органа обоняния благодаря стимуляции тока воды дыхательными движениями. В то же самое время, находясь в ротовой полости, задняя ноздря оказалась пригодной функционировать как хоана наземных позвоночных, что обеспечило проведение воздуха через нос и возможность атмосферного дыхания. Экспериментальным подтверждением происхождения хоаны из задней ноздри рыб является совместная локализация презумптивного материала ноздри и хоанального канала у амфибий соответственно единой закладке передней и задней ноздрей у рыб. Согласно гипотезе И. И. Шмальгаузена, хоана наземных позвоночных является производным задней ноздри рыб, а именно ее медиальной части, ибо предполагается, что задняя ноздря предварительно делилась на две (у некоторых кистеперых, например у Porolepis, задняя наружная ноздря сосуществовала одновременно с хоаной (Воробьева и Обручев, 1964)). Соответственно хоанальный канал должен являться производным канала задней ноздри рыб. Установление факта, что у примитивных амфибий на ранних стадиях развития закладка ноздри и ее канала располагается рядом с закладкой хоанального канала или даже является общей, как это было у рыб (хотя в сформированном органе обоняния Tetrapoda ноздря и хоана находятся на его диаметрально противоположных концах), служит еще одним доказательством происхождения хоанального канала наземных позвоночных из канала задней ноздри рыб, и, следовательно, соответственного происхождения хоаны.
Поскольку задняя и передняя ноздри перемещались к краю рта по боковой поверхности головы, а задняя ноздря затем и в глубь ротовой полости, они неизбежно должны были тянуть за собой при этом движении обонятельный мешок. Обонятельный мешок, вытянутый в ростро-каудальном направлении, испытал таким образом вращение вокруг своей продольной оси, следы которого сохранились в ходе развития якобсонова органа амфибий и его дефинитивном латеральном положении у Urodela и некоторых примитивных Anura, . То, что в филогенезе поворот обонятельного мешка амфибий действительно имел место, зафиксировано не только в эмбриональном сдвиге закладки якобсонова органа с медиальной стороны обонятельного мешка на латеральную, но и в повороте самой обонятельной плакоды (Медведева, 1965, 1968). Наличие этого поворота является еще одним доказательством правильности гипотезы И. И. Шмальгаузена о перемещении задней ноздри рыб в ротовую полость и преобразовании ее в хоану.
В главе III говорилось, что ввиду спорности и многолетнего дискути-рования вопроса о гомологии якобсонова органа у Anura и Urodela в частности и у амфибий и остальных наземных позвоночных вообще представляется недостаточным для окончательного решения проблемы ограничиться только эмбриологическими данными, показывающими общий источник возникновения этого органа. Нужно еще дать объяснение причин той огромной вариабельности в строении и положении якобсонова органа, которая существует у разных групп Tetrapoda. Филогенетический поворот обонятельного мешка, безусловно явившийся одной из причин вариабельности положения якобсонова органа, не объясняет ни наблюдаемого многообразия его строения у современных наземных позвоночных, с одной стороны, ни того обстоятельства, что, с другой стороны, этот орган имеется у всех их представителей (хотя бы в Качестве рудиментарной закладки, свидетельствующей о его полном развитии у сравнительно недавних предков). Между тем здесь имеется противоречие, заключающееся в том, что вариабельность строения указывает на наличие различных функций органа у разных групп, причем его редукция у многих форм показывает отсутствие жизненно важного значения этих функций, а существование у всех представителей наземных позвоночных хотя бы в виде рудимента свидетельствует о его прежней необходимости. Решение этого противоречия связано с вопросом о смене функций якобсонова органа в процессе исторического развития. Функции якобсонова органа у современных позвоночных до настоящего времени твердо не установлены. Большинство авторов, базируясь в основном на исследованиях функции этого органа у рептилий и млекопитающих, приходят к выводу, что он служит органом апробации пищи, находящейся в ротовой полости или в непосредственной близости от рта. Сходное мнение высказывалось и относительно амфибий (Brunei, 1914; Voss, 1936). Такая функция не делает орган жизненно необходимым, и в этом случае понятно его отсутствие у многих групп Tetrapoda.
Однако наряду с тем, что он явно не является у современных животных жизненно необходимым, его рудименты найдены у всех Tetrapoda. Это противоречие может быть объяснено только тем, что первоначальная функция якобсонова органа у исходных предков наземных позвоночных была иной и имела жизненно важное значение. Отсутствие якобсонова органа у рыб, возникновение его только у наземных позвоночных, полная или частичная редукция у вторичноводных амфибий, например у Proteidae (Seydel, 1895; Anton, 1912), Cryptobranchus (Megalobatrachus) japonicus (Fleissig, 1909), говорят о том, что этот орган не требуется для жизни в воде. С другой стороны, целый ряд вполне наземных позвоночных (птицы, млекопитающие) утратил его вторично. Напрашивается вывод, что якобсонов орган имел жизненно важную функцию, но на очень короткое историческое время, а именно он был необходим позвоночным в переходном периоде выхода из воды на сушу. Затем необходимость в нем отпала, и в различных ветвях наземных позвоночных он либо сохранился, изменив функцию, причем это изменение могло идти в разных направлениях, либо подвергся за ненадобностью редукции. Ранняя и быстрая дивергенция основных ветвей наземных позвоночных сопровождалась столь же резкой дивергенцией якобсонова органа (первоначальная функция которого оказалась ненужной), приведшей к той огромной вариабельности строения, которая так затруднила установление его гомологии.
В чем же заключалась первоначальная функция якобсонова органа? Впервые вышедшее из воды животное должно было оказаться на суше достаточно беспомощным, так как его органы чувств не были приспособлены к восприятию ощущений в воздушной среде. Это в полной мере относится и к органу обоняния. Сухая слизистая носа не могла воспринимать запахи, ибо и у наземных позвоночных воздействие пахучих веществ на обонятельные рецепторы осуществляется только после растворения в жидкой среде (Винников и Титова, 1957), а железы, характерные для наземных позвоночных и служащие для увлажнения этой слизистой, еще не появились. Однако, если у такого, впервые вышедшего из воды, животного в носовой полости имелся добавочный мешок, отходящий от дна этой полости и расположенный под нею, и к тому же снабженный обонятельным эпителием, то именно в нем могла сохраняться вода, в то время как основная часть органа обоняния сразу заполнялась воздухом (И. И. Шмальгаузен, 1964б) и, следовательно, утрачивала способность функционировать если не полностью, то в значительной мере. При этом единственным участком органа обоняния, создававшим возможность нормального функционирования обонятельных рецепторов, оставался добавочный вентральный мешок в силу сохранения в нем воды (Медведева, 1968; Jurgens, 1971). В пользу этой точки зрения говорит существование у многих рыб самых разнообразных добавочных мешков органа обоняния, из которых один, отходящий от дна обонятельной полости, мог послужить основой для якобсонова органа, и непременное, несмотря на разнообразную структуру и положение, отхождение якобсонова органа Tetrapoda от вентральной части обонятельной полости. Наличие такого мешка позволяло животному пользоваться на суше в сущности еще водным органом обоняния. Таким образом, роль якобсонова органа у переходных форм заключалась в его использовании именно в воздушной среде, хотя и как органа водного обоняния, а не в водной, как это думает Парсонс (Parsons, 1970a). В воде при наличии значительно большей поверхности, занятой сенсорным эпителием в самом обонятельном мешке, якобсонов орган мог иметь лишь подсобное значение, тогда как на суше, где слизистая обонятельного мешка не была способна полноценно функционировать без достаточного увлажнения, он оказался жизненно необходимым. С последующим возникновением в слизистой носа желез, а в дальнейшем и слезно-носового протока, обеспечивших ее увлажнение, это жизненно важное значение якобсонова органа было утрачено. Первые наземные позвоночные, расселяясь по суше, пошли по пути широкой адаптивной радиации и дивергенции. Соответственно якобсонов орган либо перестраивался в связи со сменой, функции, либо в какой-то мере сохранял первоначальную, либо вообще оказывался ненужным и редуцировался. Специфика исторического пути этого образования обусловила его огромную вариабельность у современных Tetrapoda как в отношении строения, так и в отношении функции, и тем самым затруднила решение вопроса о его гомологии.
В ходе эволюционного преобразования органа обоняния водных животных в орган обоняния наземных позвоночных произошло его объединение в единую систему с отрезком инфраорбитального сейсмосенсорного канала, превратившегося в слезно-носовой проток. Ведущим процессом в этом преобразовании было передвижение ноздрей, повлекшее за собой поворот обонятельного мешка. Опускаясь к краю рта, обе ноздри рыб пересекли линию прохождения инфраорбитального канала. И если задняя ноздря продолжала затем свой путь дальше, в ротовую полость, то передняя ноздря, как это показывает эмбриональное развитие хвостатых амфибий, вернулась вторично на дорсальную сторону головы, захватив с собой передний конец пересеченного ею отрезка инфраорбитального канала. Каудальный конец этого отрезка после ухода задней ноздри в ротовую полость заканчивался свободно в коже. В это время он еще, конечно, функционировал как сейсмосенсорный канал. Никакой связи с глазом еще не было и поэтому не могло быть непосредственного перехода к функции слезно-носового протока, заключающейся в увлажнении слизистой органа обоняния за счет выделений глазных желез. Л. П. Татаринов (1969), например, считает, что у Ichthyostega - наиболее примитивной из известных в настоящее время форм ископаемых амфибий, передняя часть слезно-носового протока, проходящая через septomaxillare, еще выполняла функцию сейсмосенсорного канала. Однако перестав функционировать как сейсмосенсорный канал, этот отрезок не был утрачен, а сохранился до своего последующего превращения в слезно-носовой проток, которое не могло быть непосредственным, так как отсутствовала связь с глазом. Следовательно, у этого отрезка инфраорбитального канала должна была быть в промежуточный период какая-то третья функция, которая помогла ему как сохраниться, так и установить связь с глазом для дальнейшего преобразования в слезно-носовой проток. Возникает вопрос: в чем могла состоять эта функция? Будем исходить из фактов. Задняя ноздря передвигается к краю рта, а затем в ротовую полость, создавая тем самым улучшенные условия обоняния для неподвижного хищника, затаившегося в засаде. Но хищник не все время сидит в засаде, он должен и передвигаться и при этом ему тоже нужно обоняние, хотя бы для того, чтобы найти партнера противоположного пола. А задняя ноздря, т. е. выводное отверстие, уже находится в ротовой полости и при движении рыбы тока воды через обонятельную полость не происходит или он сильно затруднен. Вот здесь-то и пригодилась дополнительная связь органа обоняния с внешней средой, предоставленная ему соединенным с передней ноздрей отрезком инфраорбитального канала, который мог заменить заднюю ноздрю. Однако положение вводного и выводного отверстий рядом друг с другом не благоприятствует току воды через орган обоняния, и передний конец отрезка инфраорбитального сейсмосенсорного канала начал смещаться по каналу передней ноздри к обонятельному мешку, тем самым улучшая условия смены воды в обонятельной полости. Он стал дублировать канал задней ноздри, и его дальнейшее историческое развитие пошло по тому же пути. Этот путь до сих пор воспроизводится в эмбриональном развитии примитивных амфибий передним концом слезно-носового протока и еще раз подтверждается в ходе редукции протока у Pleurodeles waltlii. Каудальный конец отделившегося отрезка инфраорбитального канала при этом сместился назад и подошел к глазу - была приобретена возможность для осуществления дефинитивной функции слезно-носового протока, которая оказалась тем более кстати, что это произошло уже у животного, периодически выходившего на сушу. Мне представляется совершенно правильным предположение И. И. Шмальгаузена об одновременном наличии у переходных форм хоаны и задней наружной ноздри, но я думаю, что их возникновение не было результатом деления задней наружной ноздри рыб на две. Просто ее функцию взял на себя соединившийся с органом обоняния отрезок инфраорбитального сейсмосенсорного канала, сделавший таким образом первый шаг на пути своего превращения в слезно-носовой проток. Третья наружная ноздря некоторых кистеперых, видимо, и является каудальным отверстием будущего слезно-носового протока. Соответственно слезно-носовой проток не имеет никакого отношения по своему происхождению к каналу задней ноздри (как это полагали Эллис (Allis, 1932a) и Ярвик (Jarvik, 1942) и что горячо поддержал в последнее время Бертмар (Bertmar, 1969)), он просто в период своего исторического становления имитировал строение и расположение этого канала вследствие исполнения сходной функции.
На основании всего изложенного можно попытаться восстановить схему строения органа обоняния предков наземных позвоночных. Предполагаемый ход эволюционных преобразований в процессе перехода позвоночных от водной к наземной жизни, показан на рис. 78. Исходным для возникновения органа обоняния наземных позвоночных должен был послужить обонятельный мешок рыб достаточно простого строения, без добавочных мешков (на что указывает нерасчлененный обонятельный мешок хвостатых амфибий и гимнофион), открывавшийся двумя наружными ноздрями - передней и задней, вытянутыми в сравнительно длинные трубки (так как иначе передвижение ноздрей вызывало бы слишком резкое изменение положения обонятельного мешка) на дорсальной стороне головы (рис. 78, А). Обонятельный эпителий сосредотачивался на вентральной стороне мешка, как это вообще характерно для рыб. Передвижение обеих ноздрей к краю рта привело к пересечению ими инфраорбитального сейсмосенсорного канала (рис. 78, Б). В результате поворота обонятельного мешка область обонятельного эпителия переместилась на медиальную сторону мешка. Именно в области, занятой обонятельным эпителием, возник в это время добавочный мешок, послуживший основой для будущего якобсонова органа, на что указывают первичность его медиальной закладки у всех наземных позвоночных и наличие в нем обонятельного эпителия, а также эмбриональное передвижение якобсонова органа современных амфибий без первоначального смещения в медиальную сторону, которое было бы неизбежным, если бы добавочный мешок, послуживший основой для якобсонова органа, образовался на более ранней стадии исторического развития, в то время как область, содержащая обонятельный эпителий, находилась на вентральной стороне органа обоняния.
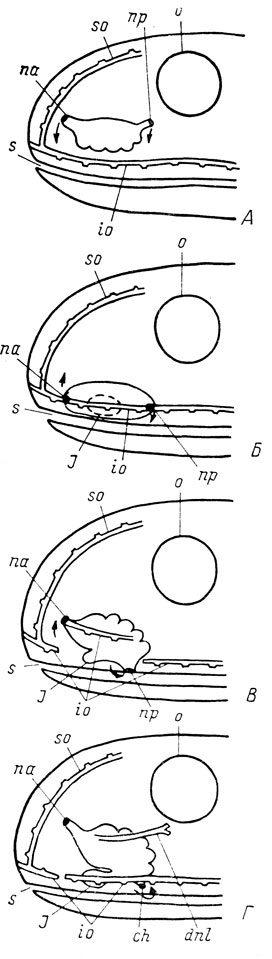
Рис. 78. Схема исторического преобразования органа обоняния позвоночных в процессе перехода от водной к наземной жизни. Стрелки показывают путь передвижения ноздрей. А - исходный тип органа обоняния водного животного; Б, Б - промежуточные стадии исторического развития; Г - тип органа обоняния примитивных наземных позвоночных
После достижения края рта задняя ноздря продолжала смещаться в ротовую полость, поскольку попадала при этом в зону отрицательного давления, возникавшую при дыхательных движениях рыбы, что способствовало лучшему просасыванию воды через орган обоняния. Если бы отрезок инфраорбитального канала, который пересекла на своем пути задняя ноздря, оставался соединенным с ней, то ему пришлось бы погрузиться в ротовую полость, что воспрепятствовало бы его функционированию в качестве сейсмосенсорного канала. Поэтому каудальный конец отрезка канала, находившегося между обеими ноздрями, остался свободным в коже. Положение задней ноздри в ротовой полости обеспечивало эффективное просасывание воды через орган обоняния при любом положении передней ноздри и сделало излишним ее локализацию у края рта. Передняя ноздря стала подниматься наверх на дорсальную сторону головы, захватив при этом с собой соединенный с ней передний конец отрезка инфраорбитального канала, которому это перемещение не препятствовало функционировать как сейсмосенсорному, поскольку оно сохраняло его поверхностное положение. Канал передней ноздри был значительно короче, чем задней (как это и сейчас имеет место у латимерии и Polypterus), поэтому передняя часть обонятельного мешка испытывала значительно более интенсивное вращение, чем задняя, соединенная с более длинным каналом задней ноздри с ее теперь уже зафиксированным положением как хоаны в крыше ротовой полости. Именно поэтому добавочный мешок, располагавшийся медиально от органа обоняния в его передней части, сдвинулся в латеральную сторону и оказался вентрально от обонятельного мешка (рис. 78, В). Отрезок инфраорбитального канала, который, вероятно, нес в это время двойную функцию: и сейсмосенсорного канала, и канала задней ноздри, уже отходил не от края передней ноздри, а был смещен, к обонятельному мешку (рис. 78, Г). Таким путем был достигнут исходный тип для органа обоняния наземных позвоночных, уже вполне пригодный для функционирования в воздушной среде (и как орган обоняния, и как часть дыхательной системы) и очень мало отличающийся по своему строению от соответствующего органа современных хвостатых амфибий. Он уже был приспособлен для проведения атмосферного воздуха, имея ноздрю и хоану, и обеспечивал возможность использования рецепторов органа обоняния в воздушной среде благодаря наличию вентрально расположенного якобсонова органа, в котором сохранялась вода, когда животное выходило на сушу. Степень продвижения отрезка инфраорбитального сейсмосенсорного канала по пути превращения в слезно-носовой проток не играла особой роли при амфибиотическом образе жизни первых осваивавших землю позвоночных. Если палеонтологам удастся когда-нибудь найти форму, переходную от водных к наземным позвоночным и являющуюся прямым предком современных Tetrapoda, у нее должен быть орган обоняния подобного типа.
Эмбриологическое исследование органа обоняния ныне живущих амфибий показывает, что несмотря на все разнообразие его отдельных структур у хвостатых, бесхвостых и безногих амфибий, приведшее к весьма пессимистическому высказыванию о наличии у всех трех отрядов земноводных разных типов носа и отсутствии доказательства вероятной связи этих типов (Parsons, 1967), любой из отделов в системе органа восходит к единому прототипу и, следовательно, удостоверяет происхождение от общих предков. Сложные процессы образования хоан, якобсонова органа, слезно-носового протока и связанных с ним покровных костей крыши черепа - lacrymale и septomaxillare - сводятся к единому источнику не только для амфибий, но и для всех наземных позвоночных. Утверждение Ярвика (Jarvik, 1972), что его теория дифилетического происхождения амфибий имеет под собой хорошо обоснованный базис, не оправдывается строением, развитием и ходом эволюционных преобразований ни одной из структур, имеющих отношение к системе органа обоняния, весомость же некоторых его обоснований, касающихся якобсонова органа (стр. 16 - 17), septomaxillare (стр. 136 - 137), происхождения слезно-носового протока (стр. 134 - 135), филогенетического перемещения ноздрей (стр. 146 - 148), показана выше. На основании исследования одной системы нельзя, конечно, делать вывода о монофилетическом происхождении Tetrapoda, но заключение о том, что орган обоняния амфибий не дает никаких оснований для возможности утверждения их полифилетического происхождения, совершенно правомочно.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'