1. Сопоставление сравнительных и экспериментальных данных по развитию органа обоняния и слезно-носового протока амфибий
Хоанальный вырост обонятельного мешка как хвостатых, так и бесхвостых амфибий развивается не из оро-назальной борозды, которой у них не существует, а из ткани самого обонятельного мешка в области переходной зоны между обонятельным и индифферентным эпителием, у каудального края ноздри. Этот факт подтверждается результатами пересадки обонятельной плакоды в область слухового пузырька, где материала оро-назальной борозды быть не может, а развившийся в этих условиях обонятельный мешок все же образует хоанальный вырост.
Однако хоанальный канал в целом развивается не только за счет хоанального выроста обонятельного мешка, но и у х:востатых в равной, а у бесхвостых амфибий в меньшей степени за счет растущего ему навстречу выроста передней кишки. Этот факт, который подчеркнул в своем исследовании Кавагое (Kawagoe, 1932), считая, что он относится только к развитию органа обоняния у японского скрытожаберника Megalobatrachus japonicus, справедлив для всех хвостатых и бесхвостых амфибий и был вскользь отмечен в свое время еще Гинсбергом (Hinsberg, 1901), но не привлек к себе должного внимания исследователей. Он полностью подтверждается тем, что при пересадке обонятельной плакоды на место слухового пузырька соединение хоанального выроста с глоткой осуществляется только при наличии растущего ему навстречу выроста эпителия глотки, а также самостоятельным образованием хоаны после полного удаления органа обоняния. Эта образующаяся в отсутствие органа обоняния хоана представляет собой значительное углубление дорсальной стенки ротовой полости, особенно у Hynobiidae, по-видимому, включающее в себя также и самую каудальную часть хоанального канала, поскольку в норме у хвостатых амфибий последняя развивается не за счет хоанального выроста обонятельного мешка, а за счет хоанального выроста стенки ротовой полости. Хотя у бесхвостых амфибий при нормальном развитии хоанальный вырост ротовой полости представляет собой всего лишь небольшой выступ, который трудно рассматривать как каудальную часть хоанального канала, наличие после полного удаления обонятельной плакоды 5 случаев с образованием не только хоаны, но и этой части хоанального канала показывает, что и у Anura, она первоначально возникала совместно с хоаной из материала передней кишки. Таким образом, следует говорить о самостоятельном образовании у амфибий не только собственно хоаны, но и прилежащей к ней части хоанального канала.
В литературе имеются только отдельные разрозненные данные о самостоятельном возникновении хоаны после удаления закладки органа обоняния. Шпеман (Spemann, 1912) наблюдал 1 случай образования хоаны у Rana esculenta при отсутствии связи обонятельной закладки с ротовой полостью. Экман (Ekman, 1923) упоминает о 3 таких же случаях у бесхвостых амфибий и на этом основании заключает, что хоана имеет энтодермальное происхождение. 5 случаев образования несовершенной хоаны у R. pipiens после полного удаления обонятельной плакоды описаны Купер (Cooper, 1943). Немногочисленность этих данных, а также отсутствие данных по хвостатым амфибиям объясняются только тем, что в отсутствие органа обоняния хоаны, в особенности у высших тритонов, образуются поздно, а исследователи, удалявшие закладку органа обоняния, занимались в основном проблемами регенерации и фиксировали материал на более ранних стадиях развития.
В вышедшей не так давно работе Тейхмана (Teichmann, 1962) разбирается вопрос о формообразовании хоан у хвостатых амфибий Triturus alpestrisn T. vulgaris (Triton taeniatus). В ней он показывает в соответствии с изложенными здесь и ранее (Медведева, 1960а, 1961а, 1964) результатами, что хоанальный канал хвостатых амфибий развивается путем соединения хоанального выроста обонятельного мешка с растущим ему навстречу выростом ротовой полости. Что же касается образования самой хоаны, то Тейхман пришел к ошибочным выводам. Он полагает, что при удалении даже не всей обонятельной плакоды, а только ее задней половины хоана вообще не развивается. Причина ошибки все та же, что и у предыдущих исследователей, - Тейхман фиксировал подопытных личинок слишком рано после времени проведения операций, а у Salamandridae, к которым относятся оба исследованных им вида, как показано в главе IV, хоана на оперированной стороне образуется значительно позже, чем в норме. Поэтому Тейхман принял за отсутствие хоаны отсутствие хоанального канала (т. е. соединения хоанального выроста обонятельного мешка с выростом ротовой полости, в котором и возникает собственно хоана), что действительно имеет место при удалении задней половины обонятельной плакоды, так как именно в ней расположен презумптивный материал хоанального канала не только у Salamandridae, но и у всех хвостатых и бесхвостых амфибий (см. главу IV). Все приведенные выше факты, а также более ранние данные того же Тейхмана (Teichmann, 1955, 1959, 1961) об образовании у Triturus alpestris при гомо- и ксенотрансплантации обонятельной закладки на различные участки головы и туловища всех частей органа обоняния кроме хоаны и соответствующие данные Купер (Cooper, 1943) по Rana pipiens показывают, что у хвостатых и бесхвостых амфибий презумптивный материал хоаны, хотя она и является неотъемлемой частью органа обоняния, не содержится в обонятельной плакоде, и полностью подтверждают вывод о самостоятельности образования собственно хоаны и ее развитии не за счет ткани обонятельного мешка, а за счет материала стенки ротовой полости.
В то же время как у хвостатых, так и у бесхвостых амфибий возникающие вне морфологической связи друге другом обонятельный мешок и хоана объединены формативной связью типа эмбриональной индукции (у бесхвостых амфибий, так же как и у высших хвостатых, хоана развивается при полном отсутствии органа обоняния, но запаздывает во времени по сравнению с нормой). Возникновение и усиление значения этой формообразовательной связи с большой ясностью прослеживается экспериментально в ряду хвостатых амфибий от низших к высшим. Допущение возможности ранней индукции (до удаления закладки органа обоняния) и предварительной детерминации места закладки хоаны не опровергает выявленных закономерностей, так как и в этом случае наличие органа обоняния остается необходимым для образования полноценной хоаны и ее сохранения у высших хвостатых амфибий и не является непременным условием у низших. Тот факт, что формообразовательная связь между органом обоняния и хоаной у хвостатых и бесхвостых амфибий находится еще в процессе исторического становления, указывает, во-первых, на то, что эта система образовалась сравнительно недавно, и, во-вторых, на то, что должны были быть предшествовавшие отношения между обонятельным мешком и хоаной, на базе которых могло произойти возникновение новой формообразовательной связи.
Все сказанное относится только к хвостатым и бесхвостым амфибиям, поскольку у гимнофион так же, как и у остальных обладающих хоанами позвоночных, хоанальный канал и хоана развиваются за счет погружения оро-назальной борозды и, следовательно, представляют, собой участки единой закладки органа обоняния.
Таким образом, среди всех современных позвоночных, обладающих хоанами, только хвостатым и бесхвостым амфибиям присущ совершенно своеобразный способ образования хоаны и хоанального канала, результатом которого является развитие органа обоняния из двух морфологически обособленных частей. С одной стороны, это обстоятельство с очень большой достоверностью указывает на то, что хвостатые и бесхвостые амфибии произошли от общих предков, уже имевших хоаны, развивавшиеся таким же путем, ибо предположение о возможности конвергентного приобретения каждой группой в отдельности столь сложного способа образования единого органа обоняния из двух совершенно отдельных, морфологически обособленных, закладок, к тому же могущих принадлежать к разным зародышевым листкам, не выдерживает критики. Даже если допустить такую возможность конвергентного (или параллельного - это зависит от того, как расценивать близость друг к другу обоих отрядов) возникновения подобного способа развития органа обоняния хвостатых и бесхвостых амфибий, то следы различного происхождения этого способа должны были бы выявиться в ходе развития органа, а как показывает вышеизложенный материал, никаких принципиальных различий в этом отношении между хвостатыми и бесхвостыми амфибиями не наблюдается. С другой стороны, предполагать, что хвостатые и бесхвостые амфибии приобрели хоаны независимо от своих ближайших родственников - гимнофион, сходных по данному признаку со всеми остальными наземными позвоночными, и, следовательно, признать тем самым совершенно различное происхождение хвостатых и бесхвостых амфибий в одном случае и гимнофион и высших Tetrapoda - в другом нет никаких достаточных оснований. Вот это, казалось бы, безвыходное положение и заставило многих исследователей так упорно искать у хвостатых и бесхвостых амфибий несуществующую у них оро-назальную борозду. Однако, как будет показано в следующем разделе, для отсутствия оро-назальной борозды именно у Anura, и Urodela есть реальное объяснение, основанное на ряде имеющихся в настоящее время сравнительно-эмбриологических и палеонтологических данных.
Первостепенное значение для этого объяснения имеет установление точной локализации в обонятельной плакоде презумптивного материала ноздри и вводного канала, с одной стороны, и хоанального канала, с другой, у низших хвостатых и низших бесхвостых амфибий. Как было отмечено выше (стр. 91 - 92), и материал ноздри с относящимся к ней вводным каналом, и материал хоанального канала располагаются у тех и других в одном и том же квадранте III обонятельной плакоды, хотя развивающиеся из этих материалов отделы дефинитивного органа обоняния находятся на его противоположных концах. Однако совместная локализация презумптивного материала ноздри и хоанального канала указывает на их исходное более тесное отношение друг к другу, вернее, на более тесное отношение тех частей органа обоняния, которые возникали из этого материала у предков амфибий. У селахий имеется только одно отверстие, соединяющее орган обоняния с внешней средой, т. е. формально только одна ноздря. Но фактически обеспечение протока воды через орган обоняния достигается благодаря удлинению этого отверстия и перекрыванию его в средней части кожными (не срастающимися друг с другом) лопастями. В результате передний участок обонятельного отверстия функционирует как передняя, а задний - как задняя ноздря, несмотря на то что само отверстие остается единым. Хотя у костных рыб имеются уже две ноздри: передняя - вводная и задняя - выводная, но закладываются они как единое отверстие, разделяющееся на два только в процессе дальнейшего эмбрионального развития срастающимися друг с другом кожными выростами краев этого отверстия, явно гомологичными таковым селахий. Уже у некоторых костных рыб, например у Polypterus и Latimeria, ноздри вытягиваются в длинные трубки, образуя таким путем вводной и выводной каналы. Все перечисленные образования (передняя ноздря, ее вводной канал, задняя ноздря, ее выводной канал) имеют таким образом общее происхождение и развиваются из единой закладки. Несмотря на это в дефинитивном органе обоняния костных рыб передняя ноздря с ее вводным каналом и задняя ноздря с ее выводным каналом находятся на противоположных концах органа. Совместное расположение презумптивного материала как ноздри с ее вводным каналом, так и хоанального канала, служащего у хвостатых и бесхвостых амфибий выводным, показывает, следовательно, происхождение последнего из канала задней ноздри костных рыб, тоже являющегося выводным. В обонятельной плакоде хвостатых и бесхвостых амфибий нет только материала самой задней ноздри, и поскольку выводное отверстие их органа обоняния - хоана - оказалось на ранних стадиях развития разобщенным с самим органом, необходимо выяснить причины этого разобщения.
Для решения этого вопроса огромное значение имеет установление точной локализации презумптивного материала самого хоанального канала и его соотношение с местоположением материала хоаны. По результатам опытов с частичным удалением обонятельной плакоды оказывается, что у низших хвостатых и низших бесхвостых амфибий презумптивный материал хоанального канала располагается в верхней части квадранта III, входящего в заднюю половину обонятельной плакоды (рис. 44), т. е. в ту ее половину, которая обращена к ротовому впячиваншо (рис. 41). Как показано настоящим исследованием, у хвостатых и бесхвостых амфибий материал хоаны находится в стенке ротовой полости. На рис. 41 видно, что между ротовым впячиванием и задней половиной обонятельной плакюды, в которой размещается презумптивный материал хоанального канала, располагается инфраорбитальная сейсмосенсорная плакода, занимающая то место, где у остальных наземных позвоночных проходит оро-назальная борозда (стр. 39 - 40), превращающаяся у них впоследствии в хоанальный канал, открывающийся хоаной в ротовую полость. Следовательно, у хвостатых и бесхвостых амфибий наблюдаются те же отношения между обонятельной плакодой и ротовым впячиванием, как и у других наземных позвоночных, за тем исключением, что у первых место прохождения оро-назальной борозды занято инфраорбитальной сейсмосенсорной плакодой.
Следующим пунктом, важным для выяснения вопроса о происхождении хоан хвостатых и бесхвостых амфибий, являются обнаруженные следы их перемещения внутри ротовой полости у вторых и следы передвижения ноздрей по наружной поверхности головы у первых. Оба эти процесса, как уже упоминалось выше (стр. 50), безусловно связаны друг с другом, а также и с наблюдающимся в течение эмбрионального развития перемещением положения якобсонова органа. Напомню, что ноздри хвостатых амфибий передвигаются из положения на конце морды назад к ротовому впячиванию, а затем, после установления связи органа обоняния с ротовой полостью путем образования хоанального канала, смещаются обратно, сначала вперед, а потом и дальше на дорсальную поверхность головы (рис. 77). Такое перемещение ноздрей прослеживается гораздо полнее и явственнее у примитивных хвостатых амфибий, чем у более высоко организованных, и это обстоятельство указывает на подобное же перемещение ноздрей в ряду их предков.
Хоаны более примитивных представителей бесхвостых амфибий - Xenopus laevis и Pelobates fuscus - закладываются в эктодермальной, наружной, части ротовой полости; хоаны других исследованных видов бесхвостых амфибий - в энтодермальной, более глубокой, ее части. Хоанальный вырост обонятельного мешка высших бесхвостых амфибий сначала подрастает к наружной, эктодермальной, части ротовой полости и лишь затем соединяется с энтодермальной, внутренней, частью. Хоана Xenopus laevis на поздних стадиях эмбрионального развития резко сдвигается из наружной части в глубь ротовой полости. Все эти факты указывают на филогенетическое перемещение хоаны из наружной части ротовой полости во внутреннюю и предшествовавшее наружное расположение хоаны, а следовательно, на происхождение хоаны от одной из двух наружных ноздрей рыб.
Резкий сдвиг хоан в глубь ротовой полости у X. laevis позволяет наглядно установить связь этого процесса с изменением положения якобсонова органа от медиального к вентральному и с поворотом обонятельного мешка (стр. 62). Поскольку же эмбриональное передвижение якобсонова органа из медиального положения в латеральном направлении наблюдается у всех амфибий, в том числе и у бесхвостых, оно удостоверяет филогенетический поворот обонятельного мешка, обусловленный перемещением хоан, исходно - ноздрей рыб, с наружной поверхности головы в глубь ротовой полости, у их общих предков. Экспериментальные данные, выявившие поворот самой обонятельной плакоды на ранних стадиях развития как у хвостатых, так и у бесхвостых амфибий, полностью согласуются с данными по развитию якобсонова органа и также подтверждают этот поворот (стр. 91). Согласование этих данных заходит удивительно далеко: у высших бесхвостых амфибий как поворот обонятельной плакоды, так и эмбриональное передвижение закладки якобсонова органа показывают, что вращение происходило сначала в одном, а затем в обратном направлении, что в точности соответствует передвижению ноздрей к краю рта и последующему возвращению в исходное положение и далее - на дорсальную сторону морды.
Весь сложный эволюционный процесс поворота обонятельного мешка, приведший к совершенно различному положению якобсонова органа у разных отрядов современных амфибий и к локализации презумптивного материала частей органа обоняния не в одинаковых участках обонятельной плакоды, сводится в результате изложенных наблюдений к единому исходному пункту - первоначальному медиальному отхождению якобсо-нова органа, что и зафиксировано у всех амфибий в одинаковом, медиальном, расположении якобсоновой железы.
Слезно-носовой проток является новоприобретением наземных позвоночных, существование его у кистеперых рыб весьма сомнительно. Однако несмотря на его органическую связь с органом обоняния в дефинитивном состоянии, по своему происхождению он не имеет отношения к этому органу. По всему ходу своей закладки и развития слезно-носовой проток - это преобразованный отрезок инфраорбитального сейсмосенсорного канала, вступивший в связь с передней ноздрей. После установления такой" связи передний конец отрезка канала смещался вглубь до обонятельного мешка, захватив с собой относящиеся к этой части инфраорбитального канала покровные кости lacrymale и septomaxillare, а каудальный его конец остался соединенным с кожей. Экспериментально доказанная индуктивная формативная связь между слёзно-носовым протоком и каналовыми костями сохранилась и доныне у некоторых современных амфибий. Не существует никаких принципиальных различий в образовании, развитии и соединении слезно-носового протока с органом обоняния у всех ныне живущих отрядов амфибий. Наличие такой одинаковой, очень сложной системы, возникшей опять-таки из двух совершенно различных источников: сейсмосенсорного канала и органа обоняния, возможно только при одном единственном условии - происхождении от общих предков, уже обладавших этой системой.
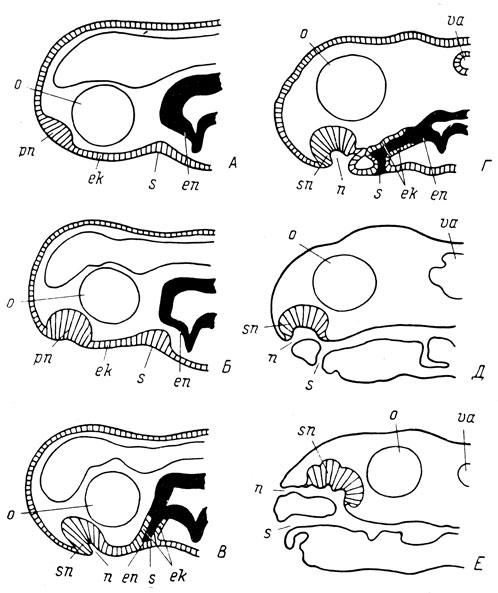
Рис. 77. Схема передвижения ноздрей и рта и изменения соотношения эктодермы и энтодермы в онтогенезе хвостатых амфибий. А - обонятельная плакода расположена на конце морды, ротового впячивания еще нет; Б - обонятельная плакода сдвинулась на вентральную поверхность морды, образующаяся обонятельная ямка направлена вентрально, ротовое впячивание эктодермальное; В - наружная часть обонятельного мешка вместе с ноздрей сдвинулась в каудальную сторону к ротовому впячиванию, ротовое впячивание экто-энтодермальное; Г - наиболее близкое расположение ноздри и ротового впячивания по отношению друг к другу в момент соединения хоанального выроста обонятельного мешка с выростом передней кишки; Д - обратное перемещение ноздри на конец морды и сдвиг ротового отверстия по вентральной поверхности морды вперед; Е - окончательное положение ноздри на дорсальной поверхности морды и конечное положение рта
В процессе эволюционного преобразования органа обоняния водных животных в орган обоняния наземных животных одновременно с морфологическими перестройками утрачивались старые и возникали новые формообразовательные связи между отдельными частями органа. С точки зрения исследования механизмов развития наибольший интерес представляет становление новых формативных зависимостей, поскольку, как показано на примере системы обонятельная закладка - хоана, подобные индуктивные аппараты возникали и формировались постепенно, совершенствуясь в ходе филогенетического развития. Для обоих, отмеченных в системе органа обоняния, формообразовательных аппаратов (обонятельная закладка - хоана и обонятельный мешок - слезно-носовой проток) характерным является первично независимое развитие составляющих компонентов. Можно полагать это общей закономерностью для систем такого рода, а их появление и дальнейшее прогрессивное развитие обусловлено наличием в них двусторонней связи между индуктором и реагирующей тканью, что обеспечивает более совершенное и правильное развитие индуцируемого органа. Заслуживает пристального внимания тот факт, что обе выявленные в настоящем исследовании вновь возникшие формообразовательные системы: обонятельная закладка - хоана и обонятельный мешок - слезно-носовой проток, составлены из компонентов, исходно имевших общее происхождение. Так, обонятельная закладка и хоана хвостатых и бесхвостых амфибий восходят к неразделенной обонятельной плакоде рыб, а обонятельный мешок и слезно-носовой проток хотя и относятся к разного типа сенсорным плакодам, но более ранний источник этих плакод единый - материал нервной пластинки. Поскольку базой для возникновения рассматриваемых формообразовательных систем послужила исторически предшествовавшая общность их компонентов, следовательно, системы такого рода могут возникать в филогенезе на основе разделения первоначально единой закладки. Предшествовавшее историческое единство индуктора и реагирующей ткани подразумевает первоначальное единство химического состава компонентов системы. Это обстоятельство может представлять очень большой интерес для решения вопроса о механизме индукционных процессов.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'