4. Обсуждение результатов экспериментального исследования развития слезно-носового протока
Экспериментальное исследование развития слезно-носового протока хвостатых амфибий и связанных с ним костей lacrymale и septomaxillare дало ряд следующих интересных результатов, хорошо согласующихся с данными нормального развития протока.
1. Доказана локализация презумптивного материала слезно-носового протока в области орбито-назального выступа инфраорбитальной сейсмосенсорной плакоды не только у низших, но и у высших хвостатых амфибий. Наряду с этим выяснилось, что тесная связь презумптивного материала слезно-носового протока с его морфологически оформленной закладкой значительно ослабевает при переходе от низших к высшим хвостатым амфибиям и одновременно повышается способность этого материала к регуляции.
2. Доказано, что развивающаяся в мезенхимной оболочке средней и дистальной частей слезно-носового протока низших хвостатых амфибий покровная кость lacrymale полностью зависит в своем образовании от протока и, следовательно, объединена с ним формообразовательной индуктивной связью, так же как каналовые кости рыб с каналами боковой линии.
3. Выявлено, что покровная кость, принимаемая у высших хвостатых амфибий за praefrontale и занимающая у них положение lacrymale и praefrontale низших хвостатых амфибий, у некоторых представителей Salamandridae, а именно у Triton cristatus karelini и Pleurodeles waltlii, показывает в своем развитии формативную индуктивную зависимость от каудальной половины слезно-носового протока, аналогичную подобной зависимости lacrymale у низших хвостатых амфибий, но не столь полно выраженную.
4. Доказано, что у низших хвостатых амфибий вторая, связанная со слезно-носовым протоком, кость septomaxillare, развивающаяся в мезенхимной оболочке его проксимального конца, также обнаруживает формативную зависимость от наличия этого конца протока и, следовательно, безусловно является по своему происхождению покровной, каналовой костью. Однако эта формативная зависимость septomaxillare от слезно-носового протока значительно ослаблена по сравнению с такой же зависимостью lacrymale и ограничивается изменением формы кости при отсутствии проксимального конца протока.
5. В случаях образования дефектного слезно-носового протока часто наблюдается его раздвоение и образование дополнительных ветвей его каудального конца, что указывает на первоначальную множественность закладок протока. На это же указывают возможность образования двух протоков: обычного и необычного (в случае удаления обонятельной плакоды), а также множественные закладки полостей во внутрикожном тяже у P. fuscus. Дополнительным доказательством исходной множественности закладок слезно-носового протока служат независимая закладка одной из двух ветвей его дистального конца (латеральной или медиальной), у хвостатых амфибий, возникновение независимой закладки третьей, добавочной, ветви, встречающееся как у хвостатых, так и у бесхвостых амфибий, а также отсутствие одной из ветвей у некоторых видов, например у Xenopus laevis (стр. 111) и Hynobius lichenatus (Watanabe, 1936b).
6. Образование необычного слезно-носового протока в случаях удаления обонятельной плакоды, его рост при отсутствии обонятельного мешка к хоане и последующее впадение в нее могут быть объяснены только наличием формативного индуктивного влияния со стороны хоаны по отношению к протоку и свидетельствуют о большой древности связи слезно-носового протока с органом обоняния, поскольку даже при отсутствии последнего проток продолжает расти в направлении морфологически отдельно развивающейся части этого органа - хоаны.
7. Обнаружено, что у примитивных хвостатых амфибий сем. Hynobiidae при отсутствии проксимального конца слезно-носового протока латеральный дивертикул обонятельного мешка, к которому в норме присоединяется этот проксимальный конец, образует воспринимающий выступ, по-видимому, представляющий собой, судя по расположению его клеток, самый проксимальный участок протока, возникающий за счет обонятельного мешка. У высших хвостатых амфибий сем. Salamandridae никаких признаков образования такого выступа не наблюдается (рис. 76).
Сопоставление результатов экспериментов и данных по нормальному развитию слезно-носового протока амфибий позволяет восстановить с достаточной ясностью картину его исторического возникновения. Локализация презумптивного материала слезно-носового протока в орбито-назальном выступе инфраорбитальной сейсмосенсорной плакоды, связь со слезно-носовым протоком покровных костей lacrymale и septomaxillare, по своему происхождению представляющих собой каналовые кости именно инфраорбитального сейсмосенсорного канала костных рыб, более того, обнаруженная формативная индуктивная зависимость развития этих костей от наличия протока с непреложностью документируют, что слезно-носовой проток наземных позвоночных является не производным органа обоняния, не новообразованием и не железистой структурой, а преобразованным отрезком инфраорбитального канала боковой линии рыб, утратившим сейсмосенсорные органы по мере приобретения этим отрезком новой функции, но сохранившим ряд вышеперечисленных признаков, характерных для каналов боковой линии. Кроме того, в развитии слезно-носового протока прослеживается исходная множественность его закладок, присущая каналам боковой линии рыб и выражающаяся в независимом развитии концевых ветвей и образовании полости протока из отдельных полостей у P. fuscus, а также в возникновении добавочных ветвей дистального конца, что напоминает ветвление наружных канальцев у кистеперых рыб. Гипотеза И. И. Шмальгаузена (1958 г) о происхождении слезно-носового протока наземных позвоночных из инфраорбитального сейсмосенсорного канала теперь может рассматриваться как теория, подтвержденная экспериментально. Следовательно, слезно-носовой проток наземных позвоночных представляет собой единственный до сих пор известный пример сохранения у них, хотя и в преобразованном виде, канала боковой линии рыб.
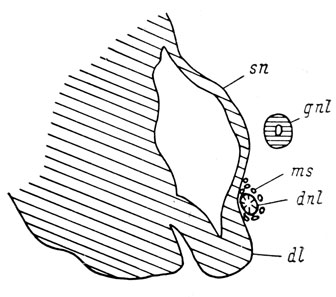
Рис. 76. Поперечный срез через латеральный дивертикул правого органа обоняния личинки Pleurodeles waltlii длиной 36 мм. Проксимальный конец слезно-носового протока достиг обонятельного мешка, стенка дивертикула никакого выступа не образует
Образование латеральным дивертикулом обонятельного мешка низших хвостатых амфибий воспринимающего выступа, который можно расценивать как самую проксимальную, хотя и очень небольшую, часть слезно-носового протока, хорошо согласуется с данными Ватанабе (Watanabe, 1936b) о наличии в носовой части протока как хвостатых, так и бесхвостых амфибий ресничного эпителия, отсутствующего на всем остальном протяжении протока, но характерного для выстилки полости обонятельного мешка, и указывает на участие в окончательном формировании слезно-носового протока также и органа обоняния. Таким образом, как и предполагал И. И. Шмальгаузен (1958 г), происхождение слезно-носового протока двойственное: несмотря на то, что в основном он представляет собой производное инфраорбитального канала органов боковой линии, его, хотя и очень незначительный, участок обязан своим существованием органу обоняния.
Какое отношение имеет этот участок слезно-носового протока, развивающийся от органа обоняния, к каналу задней ноздри рыб, от которого, как считали Эллис (Allis, 1932a) и Ярвик (Jarvik, 1942), произошел слезно-носовой проток, по имеющимся в настоящее время данным судить трудно. Развитие слезно-носового протока показывает его первоначальную, хотя и не исходную, очень древнюю связь не с задней, а с передней ноздрей. С другой стороны, в условиях эксперимента, имеющего своим результатом отсутствие органа обоняния, слезно-носовой проток растет не к передней ноздре, а к хоане, т. е. фактически к задней ноздре, причем его положение представляется инвертированным, так как на первый взгляд отходящим от кожи является его передний конец, т. е. именно тот, который в норме присоединяется к обонятельному мешку, а к хоане подрастает каудальный конец протока, который в норме отходит от покровов. Однако допустить такое необычное полное обращение концов слезно-носового протока логически невозможно, и нужно искать более обоснованную причину этому, в корне отклоняющемуся от нормы, прохождению протока.
Рост слезно-носового протока по направлению к обонятельному мешку у высших хвостатых амфибий, у которых утрачена первоначальная связь протока с ноздрей, как и рост необычного слезно-носового протока, возникающего в эксперименте, к хоане, которая представляет собой часть органа обоняния, говорит о том, что в процессе своего исторического преобразования из инфраорбитального сейсмосенсорного канала слезно-носовой проток приобрел формативную индуктивную зависимость от органа обоняния, и его развитие и направление его роста контролируются этим органом. Этой возникшей формативной зависимостью слезно-носового протока объясняется и его якобы инвертированное прохождение в экспериментальных условиях. Само образование закладки слезно-носового протока еще не зависит от наличия органа обоняния, поскольку происходит и в его отсутствие. При этом развивается только передний конец протока, который начинает расти, как и в норме, к месту, где должен располагаться обонятельный мешок, разрастания же закладки протока назад при отсутствии индуктивного воздействия со стороны органа обоняния не происходит. Поэтому поскольку при отсутствии обонятельного мешка, а следовательно и индуктивного воздействия с его стороны, передний конец слезно-носового протока изменяет свое первоначальное направление, поворачивает и начинает расти к хоане явно под индуктивным воздействием с ее стороны, этот передний конец оказывается в результате расположенным значительно более каудально, чем настоящий каудальный конец протока, остающийся связанным с кожей и лишенный возможности разрастаться назад. Хоана же, как часть органа обоняния, определенно оказывает индуктивное воздействие на погруженный в глубь тканей передний конец слезно-носового протока, результатом чего является подрастание к ней этого переднего конца и возникновение мнимой инверсии прохождения слезно-носового протока. Таким образом, соединение слезно-носового протока (при отсутствии обонятельного мешка) с хоаной не говорит о его первоначальной связи с задней ноздрей, а только подтверждает индуктивное воздействие органа обоняния в целом, включая хоану, на развитие слезно-носового протока. Ход этого развития, по имеющимся данным, не дает никаких подтверждений принадлежности незначительного участка слезно-носового протока, развивающегося от органа обоняния, к каналу задней ноздри. Продолжающееся постулирование Ярвиком (Jarvik, 1964, 1972) происхождения слезно-носового протока от канала задней ноздри кистеперых рыб и полное игнорирование экспериментальных данных, доказывающих происхождение протока от инфраорбитального сейсмосенсорного канала (Медведева, 1959, 1960б, 1960в), не может эти данные опровергнуть.
Обнаруженная у низших хвостатых амфибий сем. Hynobiidae полная зависимость образования lacrymale от наличия слезно-носового протока представляет собой единственный пример сохранения у наземных позвоночных чисто "рыбьих" отношений канала с каналовой костью. Неудивительно, что при столь полном сохранении этих отношений у низших хвостатых амфибий они в определенной мере удержались и у некоторых высших хвостатых амфибий. Однако именно факт сохранения у последних частичной формативной зависимости в развитии от слезно-носового протока покровной кости, принимаемой в настоящее время за praefrontale и располагающейся между frontale и maxillare, свидетельствует, что она в действительности таковой не является. Praefrontale по своему происхождению - каналовая кость не инфраорбитального, а супраорбитального сейсмосенсорного канала. Зависимость этой кости в развитии от слезно-носового протока, происходящего, как показано выше, от инфраорбитального сейсмосенсорного канала, необъяснима, если расценивать ее как истинную praefrontale, поскольку это положение требует допущения возникновения вторичной зависимости каналовой кости от другого канала у наземных позвоночных, у которых утрачены как каналы органов боковой линии, так и эти зависимости, за исключением единственного случая сохранения подобных отношений между отрезком инфраорбитального канала, превратившимся в слезно-носовой проток, и относящимися к нему костями.
Возможны 2 варианта происхождения "praefrontale" высших хвостатых амфибий: либо это вообще lacrymale, поскольку она обнаруживает формативную зависимость своего развития от слезно-носового протока и у ряда видов закладывается как типичная lacrymale низших хвостатых амфибий, либо это кость составная, включившая в себя обе кости - lacrymale и praefrontale. В последнем случае, поскольку эта кость у некоторых видов высших хвостатых амфибий может иногда демонстрировать полную зависимость своего образования от наличия слезно-носового протока, основную ее часть должна составлять lacrymale, а не praefrontale. Судя по развитию так называемой "praefrontale" у Ambystoma sp., где она показывает способность возникать из двух самостоятельных, не связанных друг с другом закладок (аналогичные данные, пока не опубликованные, получены и Н. С. Лебедкиной), второй вариант ближе к истине, и эта кость имеет составное происхождение и должна именоваться praefronto-lacrymale.
Таким образом, lacrymale не исчезает у большинства хвостатых амфибий, как это считалось до сих пор, а наличествует у них, хотя и в составе сложной кости, и в сохранении ее безусловно сыграло свою роль существование в преобразованном виде части инфраорбитального сейсмосенсорного канала, т. е. слезно-носового протока. Вполне возможно, что у некоторых хвостатых амфибий, у которых отсутствует слезно-носовой проток, например у японского скрытожаберника Megalobatrachus (Cryptobranchus) japonicus, именно с этим обстоятельством связано и отсутствие lacrymale. Поэтому очень интересен тот факт, что у скрытожаберника восходящий отросток maxillare, заполняя образующийся в результате отсутствия lacrymale дефект в крыше черепа, очень сильно разрастается дорсально, достигая не только praefrontale, но и frontale и оттесняя первую кость несколько назад (Wiedersheim, 1887а, 1887b), т. е. в этом случае в природе оказываются воспроизведенными фактически те же самые отношения, которые были получены в эксперименте и явились результатом отсутствия слезно-носового протока (рис. 67, Б). Можно думать, что отсутствие lacrymale у протея и амфиумы также связано у них с редукцией слезно-носового протока.
Что касается бесхвостых амфибий, то у них, как известно, отсутствуют обе кости: как lacrymale, так и praefrontale. Однако наблюдаемая в виде вариации двойная закладка сильно развитой septomaxillare у Pelobates fuscus и Rana temporaria указывает на составную природу этой кости, по крайней мере у некоторых Anura. Отдельная закладка каудальной части septomaxillare этих видов располагается в мезенхиме, окружающей среднюю часть слезно-носового протока, т. е. в положении, характерном для закладки lacrymale низших хвостатых амфибий, и лишь вторично срастается с передней и средней частями кости, представляющими истинную septomaxillare. В то же время существуют и виды бесхвостых амфибий, например Bombina bombina, у которых septomaxillare представлена только передней и средней частями, а каудальная часть кости, которая, судя по приведенным данным, является производной lacrymale, полностью отсутствует. Все же, основываясь на данных развития septomaxillare у бесхвостых амфибий, можно заключить, что у некоторых из них lacrymale тоже сохраняется, войдя в состав сложной кости, которую в этих случаях следует расценивать как septomaxillo-lacrymale.
Septomaxillare примитивных хвостатых амфибий также сохраняет формативную зависимость в развитии от слезно-носового протока, хотя и не столь полную, как lacrymale, и заключающуюся в изменении формы кости при отсутствии проксимального конца протока. Сохранение этой частичной формативной зависимости наряду с типичным одинаковым развитием septomaxillare примитивных хвостатых и всех бесхвостых амфибий (во всяком случае ее передней и средней частей у последних), заключающимся в закладке кости под проксимальным концом слезно-носового протока в окружающей его скелетогенной мезенхиме безо всякой связи с хрящом обонятельной капсулы (соединение с которым устанавливается лишь на более поздних стадиях развития и далеко не во всех случаях) и в дальнейшем разрастании кости вокруг протока, заканчивающемся включением его в костный канал, служит бесспорным доказательством принадлежности этой кости к инфраорбитальному сейсмосенсорному каналу и вполне достоверным обоснованием ее гомологии у Anura и Urodela.
Ярвик, в течение многих лет отстаивавший хрящевую природу septomaxillare хвостатых амфибий и присвоивший ей название narioidale (для отличия от septomaxillare Anurd), в последние годы под давлением фактов вынужден был отказаться от этого мнения (Jarvik, 1964, 1972). Однако он пытается спасти положение, гомологизируя эту кость с небольшой покровной косточкой поролепиформных кистеперых рыб, расположенной между fenestra exonarina anterior и fenestra exonarina posterior, названной сначала dermonarioidale (Jarvik, 1964), а затем просто narioidale (Jarvik, 1972), чем подчеркивается ее гомология с septomaxillare хвостатых амфибий. Гомологизация эта совершенно неубедительна. Она обосновывается, во-первых, положением костей, но покровные кости в процессе филогенетического развития чрезвычайно лабильны и их сходное положение без детального прослеживания хода исторических преобразований ничего не доказывает. Во-вторых, Ярвик ссылается на общее происхождение narioidale Porolepiformes и septomaxillare хвостатых амфибий, предполагая, что этим общим источником послужила оссифицированная латеральная стенка обонятельной капсулы, ставшая, вследствие общей тенденции к. ретро-грессивному развитию скелета, сначала хрящевой, а затем превратившаяся в покровную кость. Такое серьезное открытие, как превращение замещающих костей в покровные, безусловно требует более весомых обоснований, чем простое утверждение, что это могло быть.
В то же время все вышеприведенные данные, как по нормальному развитию septomaxillare у хвостатых и бесхвостых амфибий, так и экспериментальные, показывают, что аргументация Ярвика не имеет под собой фактических оснований. Septomaxillare хвостатых и бесхвостых амфибий по своему развитию, положению и происхождению - одна и та же кость, по-видимому гомологичная, как это утверждает Ярвик для бесхвостых амфибий (Jarvik, 1942), rostrale laterale остеолепиформных кистеперых рыб -кости инфраорбитального сейсмосенсорного канала, лежащей у края ноздри впереди lacrymale и имеющей у большинства из них pr. dermintermedius, заходящий внутрь обонятельной капсулы (Jarvik, 1966). Этот отросток характерен для septomaxillare всех бесхвостых амфибий и обнаружен И. И. Шмальгаузеном (1958 г) также у Onychodactylus fischeri, представителя наиболее примитивного семейства хвостатых амфибий - Hynobiidae, что служит дополнительным доказательством гомологии этой кости у Anura, и Urodela. Согласен с гомологией septomaxillare бесхвостых амфибий и rostrale laterale Osteolepiformes и Панчен (Panchen, 1967), видящий только затруднения в гомологизации с ними septomaxillare хвостатых амфибий. Однако эти затруднения снимаются настоящим исследованием и предыдущими (Медведева, 1959, 1960б, 1960в), дающими экспериментальные доказательства того, что septomaxillare у последних представляет собой каналовую кость переднего отрезка инфраорбитального сейсмосенсорного канала (каковой она и являлась у Osteolepiformes), превратившегося у наземных позвоночных в слезно-носовой проток.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'