2. Удаление орбито-назального выступа инфраорбитальной сейсмосенсорной плакоды у Salamandridae
Triton taenicitus. Удаление орбито-назального выступа инфра-орбитальной сейсмосенсорной плакоды производилось у эмбрионов обыкновенного тритона на стадиях 29 - 35 по Глезнеру. Всего было прооперировано, выращено и исследовано 59 экз. Фиксация проводилась начиная со стадии 59 по Глюкзон (появление первой морфологически отличимой закладки слезно-носового протока) и до конца метаморфоза. Несмотря на такие сроки фиксации материала, охватывающие весь период развития слезно-носового протока, все же в 1 случае (стадия 60) закладка протока отсутствовала на обеих сторонах.
Из оставшихся 58 личинок у одной слезно-носовой проток на оперированной стороне отсутствовал полностью и в 13 случаях он был дефектным и значительно отставал в развитии. Дефектность протока в основном осуществлялась за счет отсутствия или неправильного развития его проксимального (переднего) конца - 8 случаев. В 4 из этих случаев передний конец протока был раздвоен и либо обоими концами присоединялся к обонятельному мешку, либо дополнительный конец самостоятельно оканчивался в коже. Дефектность дистального конца слезно-носового протока наблюдалась только в 1 случае и заключалась в образовании третьей, дополнительной, ветви, чего в норме у T. taeniatus не отмечается. В 1 случае проток был сильно уменьшен в размере и недоразвит. В 3 случаях отсутствовала медиальная ветвь дистального конца протока. В норме из значительно большего количества просмотренных личинок такой случай встретился всего 1 раз, и поэтому увеличение частоты отсутствия медиальной ветви также следует отнести к признакам дефектности слезно-носового протока. В остальных 44 случаях проток был развит нормально.
Таким образом, эксперименты по удалению области презумптивного материала слезно-носового протока у T. taeniatus показывают, что у этого представителя высших хвостатых амфибий еще существует интимная связь между презумптивным материалом, локализующимся в орбито-назальном выступе инфраорбитальной сейсмосенсорной плакоды, и самим протоком, поскольку в результате получены случаи отсутствия и дефектности протока. Однако количество таких случаев много меньше, чем у Нуnobiidae, и, следовательно, эта связь значительно ослаблена по сравнению с примитивными хвостатыми амфибиями, а сам материал демонстрирует гораздо более высокую способность к регуляции. Что же касается "praefrontale", то как в случае полного отсутствия слезно-носового протока, так и в случаях его дефектности эта покровная кость была развита полностью (рис. 72). Отсюда следует, что у обыкновенного тритона не обнаруживается никаких следов формативной зависимости образования "praefrontale" от слезно-носового протока.
Рис. 72. Поперечный срез через голову личинки Triton taeniatus длиной 34 мм на уровне середины обонятельного мешка. Слева - оперированная сторона
Triton cristatus karelini. Экстирпация орбито-назального выступа инфраорбитальной сейсмосенсорной плакоды производилась у эмбрионов T. cristatus karelini на тех же самых стадиях развития, что и у эмбрионов обыкновенного тритона. Всего было прооперировано, выращено и исследовано 70 личинок. Фиксация подопытного материала проводилась в сроки образования и развития слезно-носового протока, т. е. начиная со стадии 59 по Глюкзон и до конца метаморфоза. Все же у 1 экземпляра, зафиксированного на стадии 61, закладка слезно-носового протока еще отсутствовала на обеих сторонах.
Из оставшихся 69 личинок у 53 развился нормальный слезно-носовой проток, у 2 он отсутствовал полностью и у 14 либо был дефектным, либо значительно отставал в развитии. Дефектность слезно-носового протока выражалась в отсутствии его передней (проксимальной) половины (2 случая), отсутствии каудальной половины (1 случай), раздвоении протока, при котором дополнительная часть либо тоже впадает в обонятельный мешок, либо оканчивается в коже (6 случаев), увеличении числа ветвей каудального конца до четырех - (1 случай) и в общем укорочении протока, сочетающемся с недоразвитием (3 случая). Отсутствие одной из ветвей слезно-носового протока и образование третьей, дополнительной, ветви не расценивалось как дефект, потому что такие отклонения встречаются у T. cristatus karelini и при нормальном развитии (стр. 106). В подавляющем большинстве случаев дефектности слезно-носового протока "praefrontale" была нормально развита, однако в 2 случаях полного отсутствия протока и в 1 случае отсутствия его каудальной (дистальной) половины дело обстояло по-иному. Во всех этих 3 случаях не было в наличии средней части слезно-носового протока, т. е. той части, в мезенхимной оболочке которой закладывается у низших хвостатых амфибий lacrymale. И именно в этих 3 случаях отсутствия средней части слезно-носового протока у T. cristatus karelini отсутствовала кость, принимаемая у высших хвостатых амфибий за praefrontale, т. е. за кость, являющуюся по своему происхождению каналовой костью супраорбитальной сейсмосенсорной линии. В 2 из этих случаев образовавшийся дефект в боковой части крыши черепа восполнялся разрастанием медиального выступа восходящего отростка maxillare (полное отсутствие слезно-носового протока), а в случае отсутствия каудальной половины протока этот дефект компенсировался разрастанием каудально и вентрально латерального края nasale.
Таким образом, у T. cristatus karelini не только сохранилась в определенной мере интимная связь презумптивного материала слезно-носового протока, локализованного в орбито-назальном выступе инфраорбитальной сейсмосенсорной плакоды, и самого протока, что доказывается наличием случаев с его отсутствием и дефектностью в результате удаления этого материала, но в отличие от отношений, наблюдающихся у обыкновенного тритона, выявляются и признаки еще существующей формативной зависимости между средней частью протока и покровной костью высших хвостатых амфибий, расцениваемой в настоящее время как praefrontale.
Pleurodeles waltlii. Экстирпация орбито-назального выступа инфраорбитальной сейсмосенсорной плакоды производилась у эмбрионов P. waltlii , на стадиях 27 - 33 по Гальену и Дюроше. Всего было прооперировано, выращено и исследовано 95 личинок. Хотя фиксация подопытного материала производилась в сроки образования и развития слезно-носового протока, начиная со стадии 52 и кончая завершением метаморфоза, все же поскольку развитие протока у P. waltlii очень варьирует по времени (стр. 107), 22 личинки были зафиксированы, когда закладка протока еще отсутствовала на обеих сторонах (стадии 52 - 54).
Прежде чем излагать полученные результаты, необходимо подчеркнуть, что экспериментальный материал по P. waltlii не допускает столь достоверного анализа, как соответствующий материал по другим исследованным видам, во-первых, ввиду только что отмеченной большой вариабельности в сроках закладки и развития слезно-носового протока, и, во-вторых, ввиду наличия редукции протока во второй половине метаморфоза, которая могла симулировать дефектность его в результате эксперимента. Поэтому количественное распределение различных результатов экспериментов у P. waltlii может иметь только относительное, а не абсолютное значение (при сравнении с соответствующими результатами у других представителей Salamandridae). Абсолютную же значимость безусловно имеют данные по соотносительному развитию слезно-носового протока и "praefrontale", поскольку в случае отсутствия или дефектности этой кости для делаемых выводов совершенно неважно, отсутствует (или дефектен) слезно-носовой проток в результате эксперимента или естественного процесса редукции.
За исключением 22 экземпляров, зафиксированных до начала образования слезно-носового протока, оставшиеся 73 личинки распределялись по полученным результатам следующим образом. Нормальный слезно-носовой проток развился у 24 экз. У 23 личинок он полностью отсутствовал, из них у 8 можно подозревать редукцию протока, так как она одновременно имела место на неоперированной стороне. В 26 случаях слезно-носовой проток был дефектным или значительно отставал в развитии от протока неоперированной стороны, из них в 9 случаях возможна редукция. Дефектность слезно-носового протока выражалась в общем укорочении его длины, уменьшении диаметра, отсутствии или дефектности передней (проксимальной) половины, отсутствии или дефектности каудальной (дистальной) половины.
В 1 случае отсутствия слезно-носового протока не было вообще "praefrontale" (рис. 73), в 7 случаях отсутствия протока "praefrontale" отставала в развитии или была дефектной. При дефектности слезно-носового протока случаев с отсутствием "praefrontale" не имелось, но оказалось 7 случаев, в которых эта кость значительно отставала в развитии от соответствующей кости неоперированной стороны или была дефектной (рис. 74). Все эти случаи связаны с дефектностью развития каудальной половины слезно-носового протока.
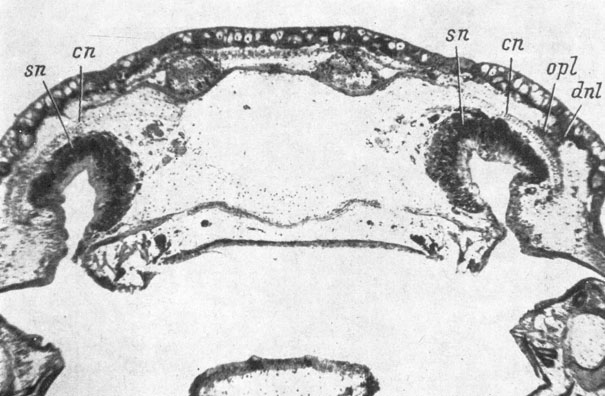
Рис. 73. Поперечный срез через голову личинки Pleurodeles waltlii длиной 58.5 мм на уровне закладки praefronto-lacrymale. (Увел. 34). Слева - оперированная сторона (нет слезно-носового протока и нет praefronto-lacrymale)
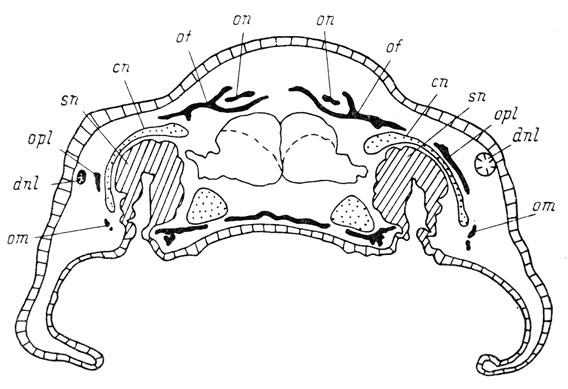
Рис. 74. Поперечный срез через голову личинки Pleurodeles waltlii длиной 38 мм на уровне закладки praefronto-lacrymale. Слева, на оперированной стороне, дефектный слезно-носовой проток и дефектная praefronto-lacrymale
Большее число случаев с отсутствием и дефектностью слезно-носового протока у P. waltlii по сравнению с количеством таких случаев у Triton taeniatus и T. cristatus karelini нельзя расценивать как доказательство сохранения более тесной связи протока с его презумптивным материалом у этого вида, так как это увеличение в данном случае могло быть спровоцировано, как указывалось выше, особенностями развития и последующей редукцией протока. Однако само наличие такой связи у Pleurodeles waltlii не вызывает сомнения, поскольку оно выявилось при соответствующих экспериментах и у других представителей Salainandridae, да и само это увеличение числа случаев свидетельствует в пользу наличия такой связи, ибо столь большое количество случаев с отсутствием и дефектностью слезно-носового протока нельзя отнести только за счет сдвига в сроках развития протока и его редукции. Что же касается соотношений в развитии слезно-носового протока и кости, принимаемой у большинства хвостатых амфибий за "praefrontale", то эта кость явно показывает сохранившуюся частичную формативную зависимость от дистальной половины протока, идентичную зависимости, выявленной у T. cristatus karelini.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'