1. Удаление орбито-назального выступа инфраорбитальной сеисмосенсорнои плакоды у Hynobiidae
Ranodon sibiricus. В первую очередь материалом для экспериментов были избраны эмбрионы R. sibiricus - реликтового вида, у которого сохранился целый ряд примитивных признаков, очень важных для целей настоящего исследования: хорошо выраженная связь слезно-носового протока с ноздрей, наличие обеих связанных с протоком покровных костей - lacrymale и septomaxillare, а также примитивность первичного расположения сейсмосенсорных линий головы (И. И. Шмальгаузен, 1955в; Медведева, 1961б). Производилось удаление презумптивного материала слезно-носового протока в месте его предполагаемой локализации - в области орбито-назального выступа (flexura lacrymalis) инфраорбитальной сеисмосенсорнои плакоды в момент образования этого выступа (эмбрионы длиной 12 - 15 мм, рис. 65). Нужно пояснить, что на стадии, на которой производились операции, никакой закладки слезно-носового протока еще не существует, так как ее первая морфологическая дифференцировка наступает значительно позднее - через 1 - 1.5 месяца после срока операции (у личинок длиной 24 - 28.5 мм), так что в данном случае речь идет лишь о возможном материале будущей закладки слезно-носового протока.
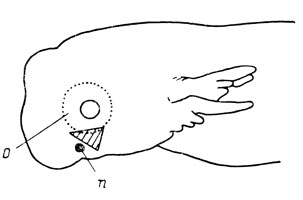
Рис. 65. Схема удаления орбито-назального выступа инфраорбитальной сеисмосенсорнои плакоды у хвостатых амфибий. Область экстирпации заштрихована
Всего было прооперировано, выращено и исследовано 45 эмбрионов. Фиксация производилась в различные сроки для получения полной картины развития слезно-носового протока в экспериментальных условиях. Последние экземпляры зафиксированы во второй половине метаморфоза, через 192 дня после операции. Поскольку полного материала по развитию R. sibiricus к моменту проведения экспериментов в руках исследователей не было, точное время закладки слезно-носового протока не было известно, и 11 личинок оказались зафиксированными на слишком ранних стадиях.
Из оставшихся 34 личинок у трех на оперированной стороне развился нормальный проток, что, по-видимому, говорит о неполноте удаления и последующей регуляции материала. Нужно отметить, что одна из этих личинок была зафиксирована на стадии первоначальной дифференцировки закладки протока, когда последняя представляет собой лишь незначительное утолщение покровного эпителия, подостланное скоплением мезенхимных клеток, так что сам слезно-носовой проток при дальнейшем развитии, возможно, мог бы иметь отклонения от нормы. Во всех остальных 6 случаях фиксации на той же стадии развития, на которой уже должна быть закладка слезно-носового протока, никаких следов этой закладки на оперированной стороне не было, что говорит либо об отставании развития протока в результате операции, либо о действительном отсутствии его, но поскольку первая возможность должна быть принята во внимание (хотя это тоже показатель локализации презумптивного материала протока в удалявшейся области), приплюсовывать эти случаи к имеющимся случаям полного отсутствия слезно-носового протока (при фиксации на более поздних стадиях развития) нельзя. Таких случаев полного отсутствия слезно-носового протока среди личинок, зафиксированных на более поздних стадиях (последние экземпляры - во второй половине метаморфоза), имеется 5. Следовательно, презумптивный материал слезно-носового протока действительно располагается в области орбито-назального выступа инфраорбитальной сейсмосенсорной плакоды.
В остальных 20 случаях на оперированной стороне образовался слезно-носовой проток, но он оказался дефектным (укорочение общей длины, уменьшение диаметра, отсутствие латеральной ветви или вообще разветвления дистального конца, недоразвитие проксимального или дистального концов) и заметно отставал в развитии по сравнению с протоком неоперированной стороны. Все эти явления говорят о нехватке материала для построения слезно-носового протока и наряду со случаями его полного отсутствия подтверждают расположение этого материала в удалявшемся участке инфраорбитальной сейсмосенсорной плакоды.
Необходимо отметить, что самая проксимальная, очень небольшая, часть слезно-носового протока, располагающаяся у места впадения его в латеральный дивертикул обонятельного мешка, может образовываться при полном отсутствии самого протока (1 случай), а также закладываться независимо в виде дорсального выступа наружной стенки латерального дивертикула обонятельного мешка (рис. 66) в тех случаях, когда проток недоразвит и вместо срастания с латеральным дивертикулом присоединяется к дорсальной (а не вентро-латеральной, как в норме на более ранних стадиях развития) стенке вводного канала (5 случаев). В 4 из 5 случаев полного отсутствия слезно-носового протока этой самой проксимальной его части в виде дорсального выступа латерального дивертикула нет, но в 3 из них фиксация была произведена как раз на стадии присоединения протока к латеральному дивертикулу обонятельного мешка, т. е. на стадии образования этого выступа (личинки длиной 45 - 50 мм), а на оперированной стороне вообще отмечается отставание развития, следовательно, выступ латерального дивертикула, представляющий собой самую проксимальную часть протока, на более поздних стадиях развития мог бы и образоваться. В последнем из случаев с полным отсутствием слезно-носового протока, когда личинка была зафиксирована во время метаморфоза, выраженного дорсального выступа латерального дивертикула тоже нет, но тут нужно учесть возможность его редукции в связи с отсутствием всего слезно-носового протока, тем более что septomaxillare в этом случае есть. Таким образом, имеющиеся случаи отсутствия дорсального выступа латерального дивертикула обонятельного мешка при отсутствии слезно-носового протока не противоречат фактам независимого образования этого выступа.
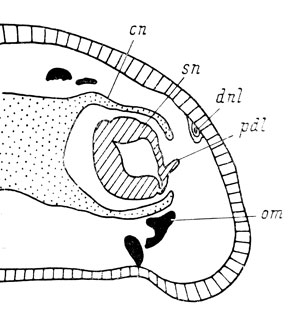
Рис. 66. Поперечный срез правого органа обоняния личинки Ranodon sibiricus длиной 60 мм на уровне присоединения слезно-носового протока к обонятельному мешку. Передняя часть протока в результате операции отсутствует, стенка обонятельного мешка образует вырост, являющийся самой проксимальной частью протока
Одним из частых дефектов развития слезно-носового протока в эксперименте является отсутствие латеральной ветви его дистального конца. Отсутствия медиальной ветви при наличии латеральной не наблюдалось ни разу. Этот факт наряду с имеющими место случаями независимой закладки латеральной ветви и ее хотя и более редко встречающимся отсутствием в норме (стр. 100) говорит о том, что медиальная ветвь, по всей вероятности, представляет собой непосредственное продолжение основного ствола слезно-носового протока, а латеральная - производное отдельной закладки.
Lacrymale закладывается в соединительнотканной оболочке средней части слезно-носового протока у личинок длиной 48 - 50 мм. Во всех 5 случаях отсутствия слезно-носового протока (при фиксации на этой и более поздних стадиях развития) lacrymale также полностью отсутствует. У личинок, зафиксированных во второй половине метаморфоза, образующийся из-за отсутствия lacrymale дефект боковой части крыши черепа восполняется за счет увеличения восходящего отростка maxillare, который разрастается дорсально и налегает верхним концом на praefrontale (рис. 67, Б). В 18 из 20 случаев образования дефектного, отстающего в развитии, слезно-носового протока lacrymale либо не образуется совсем (7 случаев), либо оказывается значительно недоразвитой (11 случаев) по сравнению с той же костью неоперированной стороны (оставшиеся 2 случая не могут быть приняты во внимание, так как у этих личинок на неоперированной стороне lacrymale еще нет). В табл. 9 приведены данные, показывающие соотношение в развитии слезно-носового протока и lacrymale на оперированной стороне тех личинок, у которых на неоперированной стороне эта кость уже образовалась.
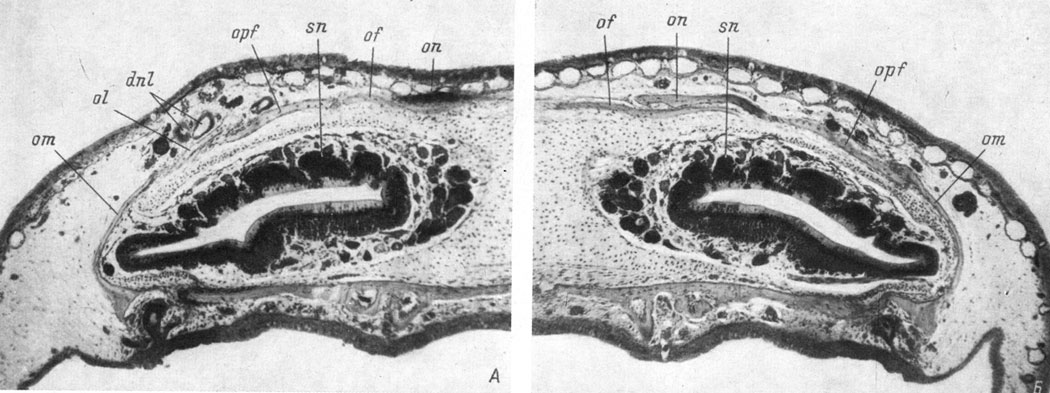
Рис. 67. Поперечный срез головы Ranodon sibiricus длиной 86 мм. (Увел. 40). А - левая контрольная сторона, есть слезно-носовой проток и есть lacrymale; Б - правая оперированная сторона, нет слезно-носового протока и нет lacrymale
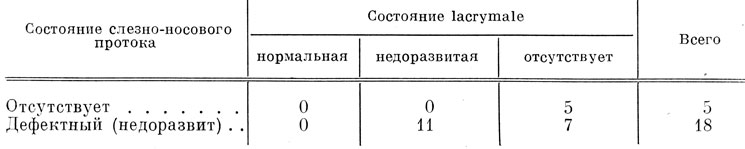
Таблица 9. Соотношение состояний слезно-носового протока и lacrymale в результате удаления орбито-назального выступа инфраорбитальной сейсмосенсорной плакоды у Ranodon sibiricus
Таким образом, lacrymale у R. sibiricus не только тесно связана в своем развитии со слезно-носовым протоком, но ее образование полностью от него зависит, поскольку во всех случаях отсутствия или дефектности протока она тоже отсутствует или является дефектной.
Septomaxillare у R. sibiricus закладывается в норме значительно позже, чем lacrymale - перед самым метаморфозом или, чаще, во время него. Разрыв в сроках закладки у различных личинок может быть очень большим. На поздних стадиях развития (во время метаморфоза), когда septomaxillare в норме уже имеется, было зафиксировано только несколько личинок, поэтому материал для полного разрешения вопроса оказался недостаточным, однако некоторые определенные результаты все же получены. У 2 личинок, зафиксированных во второй половине метаморфоза, слезно-носовой проток отсутствует полностью, а septomaxillare в обоих случаях оказывается развитой, но вместо обычной для нее кольцеобразной формы имеет вид сплошного круглого костного узелка. На рис. 68 показана закладка septomaxillare в норме, при наличии слезно-носового протока (4), и при его отсутствии в результате удаления орбито-назального выступа инфраорбитальной сейсмосенсорной плакоды (Б). В норме septomaxillare окружает основание слезно-носового протока у места его впадения в латеральный дивертикул обонятельного мешка, при отсутствии протока она остается под выступом латерального дивертикула, представляющим собой самую проксимальную часть протока, развивающуюся за счет обонятельного мешка. Этот дорсальный выступ латерального дивертикула обонятельного мешка имеется у одной из личинок с образованием septomaxillare, у другой - его наличие сомнительно, но можно, как указывалось выше, предполагать его редукцию.
Таким образом, результаты экспериментов по удалению орбито-назального выступа инфраорбитальной сейсмосенсорной плакоды у R. sibiricus показывают следующее.
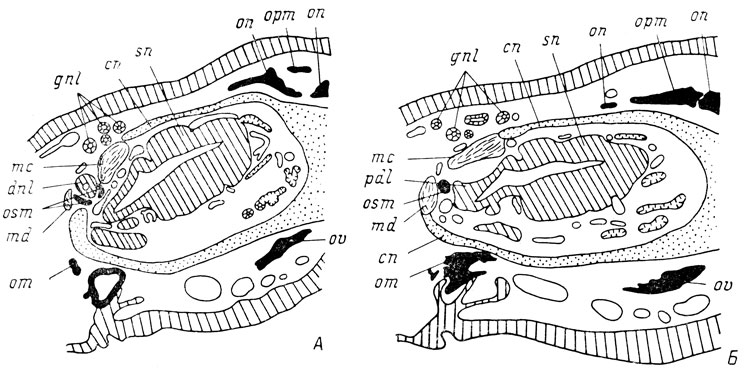
Рис. 68. Поперечные срезы головы личинок Ranodon sibiricus длиной 80 мм (А - слезно-носовой проток есть) и 83 мм (Б - слезно-носового протока нет) на уровне проксимального конца протока
1. В этой области содержится презумптивный материал практически всего слезно-носового протока, за исключением самой проксимальной и очень небольшой его части, представленной выступом латерального дивертикула обонятельного мешка.
2. Lacrymale не только образуется в мезенхиматозной муфте, окружающей слезно-носовой проток, но и полностью от него зависит в своем образовании.
3. Зависимость septomaxillare от слезно-носового протока неполная, так как эта кость может образовываться и при отсутствии протока, но все же он оказывает на нее определенное формативное воздействие, поскольку при отсутствии протока кость не может принимать присущую ей кольцеобразную форму и разрастаться дорсально от места своей закладки.
Hynobius keyserlingii. Операции заключались в удалении эктодермы в области орбито-назального выступа инфраорбитальной сейсмосенсорной плакоды у эмбрионов H. keyserlingii длиной 7 - 9.5 мм на стадиях образования первичных и вторичных жаберных выступов. Всего выжило, выращено и зафиксировано на различных стадиях развития (вплоть до конца метаморфоза) 54 личинки. Из них у 4 экз. слезно-носовой проток отсутствовал полностью, у 33 образовался в той или иной степени дефектный проток, и в 17 случаях развился нормальный проток. Такое значительное количество личинок с образованием нормального слезно-носового протока по сравнению с результатами подобных операций у Ranodon sibiricus объясняется тем, что у H. keyserlingii обонятельный мешок расположен очень близко от глаза (этим обусловлена и относительно меньшая общая длина протока), и из-за опасения повреждения обонятельного мешка (в нескольких случаях он все же был задет) презумптивный материал протока, вероятно, не был удален полностью, причем произошла его последующая регуляция.
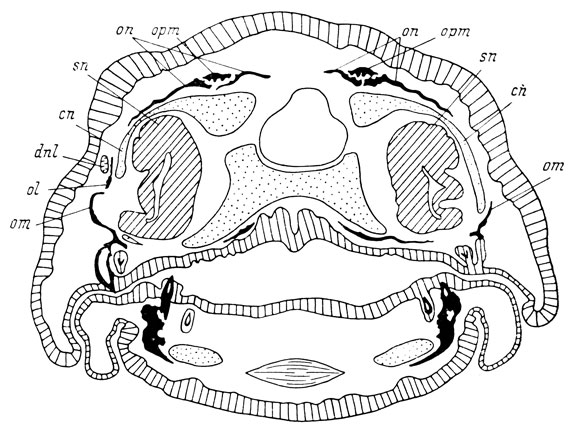
Рис. 69. Поперечный срез головы личинки Hynobius keyserlingii перед метаморфозом длиной 32 мм. На оперированной стороне (справа) нет слезно-носового протока и нет lacrymale
Из 4 случаев полного отсутствия слезно-носового протока только в одном случае фиксация была произведена на стадии, когда lacrymale в норме уже имеется, и в этом случае на оперированной стороне кости нет (рис. 69). Кроме того, lacrymale полностью отсутствовала также в 5 случаях образования дефектного и отстающего в развитии слезно-носового протока (во всех этих случаях проток деформирован очень сильно, в основном за счет отсутствия средней и дистальной частей). На поздних стадиях развития (конец метаморфоза) образующийся за счет отсутствия lacrymale дефект в боковой части крыши черепа восполняется восходящим отростком maxillare, который разрастается дорсально совершенно так же, как у Ranodon sibiricus (стр. 122). В остальных 28 случаях с образованием дефектного слезно-носового протока, за исключением одного, lacrymale также оказалась дефектной и недостаточно развитой. В 1 случае, явившемся исключением, фиксация была произведена на стадии первоначальной закладки lacrymale, так что, естественно, по нему нельзя судить о правильности развития кости. Таким образом, lacrymale показывает полную зависимость в своем развитии от слезно-носового протока, совершенно идентичную обнаруженной у R. sibiricus, что и демонстрируется в табл. 10.
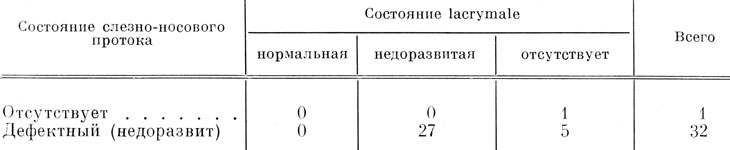
Таблица 10. Соотношение состояний слезно-носового протока и lacrymale в результате удаления орбито-назального выступа инфраорбитальной сейсмосенсорной плакоды у Hynobius keyserlingii
Необходимо отметить 2 интересных случая дефектности кости, когда слезно-носовой проток представлен только небольшим выростом, отходящим от кожи. В норме lacrymale представляет собой как описывалось выше (глава V), короткую костную трубочку, медиальная сторона которой, после окончания протока в коже, продолжается дорсо-каудально в виде пластинки, налегающей своим дорсальным концом на praefrontale. Трубочка закладывается вокруг протока в окружающей его мезенхимной оболочке, что типично для каналовых костей, и впоследствии, разрастаясь от медиальной стенки трубочки, образуется пластинка. В обсуждаемых случаях образовалась только пластинка, и этот факт представляет чрезвычайный интерес по двум причинам: во-первых, потому, что закладка каналовых костей рыб также состоит из двух частей - трубочки и пластинки (дермо-компонент и мембрано-компонент), в данных же случаях пластинка образуется не в связи с мезенхимной оболочкой слезно-носового протока, а в отдельном мезенхимном скоплении, лежащем вдоль хряща обонятельной капсулы, очень сходно с образованием мембрано-компонента некоторых покровных костей рыб; во-вторых, потому, что образование пластинки имеет место только в том случае, если сохранился хотя бы маленький отрезок дистальной части протока. Это указывает на индуктивную природу влияния слезно-носового протока на образование кости в целом, включая ее и дермо-, и мембрано-компонент.
Septomaxillare не показывает такой полной зависимости в своем развитии от наличия слезно-носового протока, как lacrymale, поскольку может образовываться как в случае полного отсутствия протока, так и в случае его дефектного развития. Причем дефектность дистальной половины слезно-носового протока не оказывает никакого влияния на развитие кости - в этом случае она развивается совершенно нормально, образуя короткую трубочку, или кольцо, вокруг имеющегося проксимального конца протока (рис. 70, А). При отсутствии же слезно-носового протока или дефектности его проксимальной половины septomaxillare развивается, но образует неправильной формы плотное костное разрастание (4 случая, рис. 70, Б). Неверно было бы усматривать в приобретении костью формы трубочки одно лишь чисто механическое воздействие на нее слезно-носового протока из-за того, что он возникает раньше, а кость, закладываясь в мезенхимной "муфте" протока, впоследствии разрастается и образует вокруг него полное кольцо. Даже при полном отсутствии слезно-носового протока можно точно определить место впадения его проксимального конца в обонятельный мешок, так как на латеральном дивертикуле независимо от наличия протока образуется воспринимающий выступ, так же как это имеет место у Ranodon sibiricus. В норме septomaxillare первоначально закладывается под слезно-носовым протоком, а затем разрастается вокруг него, охватывая основание протока со всех сторон, в том числе и с дорсальной. В случае же отсутствия протока или его проксимального конца кость также закладывается вентральнее воспринимающего выступа латерального дивертикула обонятельного мешка, но не разрастается дорсально, а образует плотное костное разрастание типа узелка на месте закладки (рис. 70, Б). То же самое отмечалось и у R. sibiricus (см. стр. 123, рис. 68, Б). Поэтому в данном случае также можно говорить об определенном формативном влиянии слезно-носового протока на развитие septomaxillare.
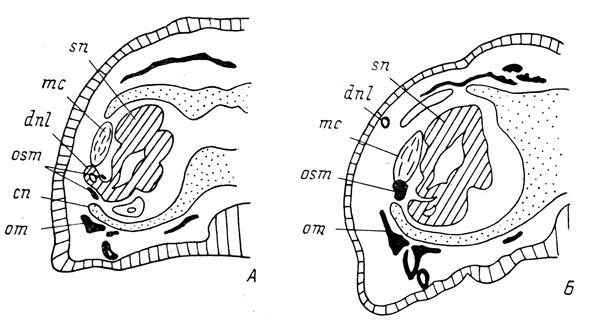
Рис. 70. Поперечные срезы через оперированную сторону головы личинок Hynobius keyserlingii длиной 34 мм (А) и 33 мм (Б). А - проксимальный конец слезно-носового протока есть, и septomaxillare образует кольцо вокруг него; Б - проксимальный конец протока отсутствует, septomaxillare - в виде плотного костного комка
У Hynobiuskeyserlingii в случае отсутствия слезно-носового протока или отсутствия соединения его с обонятельным мешком не только образуется независимый от развития самого протока выступ латерального дивертикула, но и вся вентро-латеральная стенка последнего приобретает иной характер вследствие расположения составляющих ее клеток правильными рядами и представляет собой как бы продолжение стенки самого протока (рис. 71). Этот образующийся независимо от самого слезно-носового протока выступ латерального дивертикула обонятельного мешка развился во всех случаях отсутствия соединения протока с дивертикулом, за исключением тех, в которых обонятельный мешок был поврежден. Имеется 1 случай (личинка, зафиксированная в конце метаморфоза), когда наличие выступа сомнительно, но здесь, как и в аналогичном случае у Ranodon sibiricus, можно подозревать его редукцию. Кроме того, во всех этих случаях отсутствия выступа латерального дивертикула и соответственно поврежденного и дегенерирующего обонятельного мешка отсутствует полностью также и septomaxillare, что наводит на мысль о возникновении вторичной зависимости закладки этой кости от органа обоняния. В этом случае становится понятным факт возникновения у Hynobiidae, хотя и дефектной, septomaxillare при полном отсутствии слезно-носового протока, а также описанное Ивро (Yvroud, 1971a) образование этой кости вокруг средней полости обонятельного мешка, а не вокруг переднего конца слезно-носового протока при их экспериментальном разъединении у Discoglossus pictus (Anura).
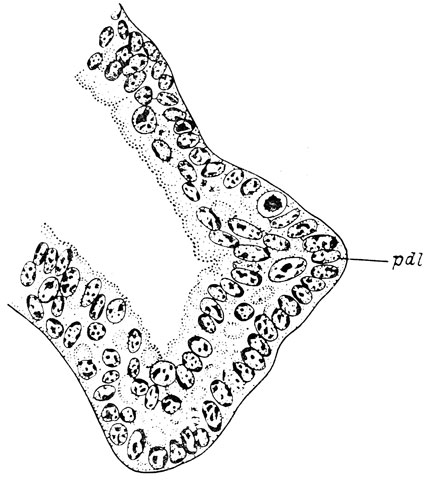
Рис. 71. Поперечный срез через латеральный дивертикул обонятельного мешка личинки Hynobius keyserlingii длиной 28 мм. Выступ дивертикула образовался независимо от слезно-носового протока, который отсутствует. Видна двухрядная ориентация клеток вентро-латеральной стенки
Таким образом, результаты исследования связи развивающегося слезно-носового протока с покровными костями lacrymale и septomaxillare у Hynobius keyserlingii полностью согласуются с результатами, полученными у Ranodon sibiricus, и дополняют друг друга. На основании сравнения и суммирования этих данных можно утверждать, что у представителей самого примитивного семейства хвостатых амфибий - Hynobiidae - сохранилась формативная зависимость покровных костей lacrymale и septomaxillare от слезно-носового протока, характерная для каналовых костей костных рыб: для lacrymale - полностью, для septomaxillare - частично. Причем образование lacrymale связано со средней и дистальной частями слезно-носового протока, a septomaxillare - с его проксимальной частью.
Эта зависимость, а также локализация презумптивного материала слезно-носового протока в области орбито-назального выступа инфраорбитальной сейсмосенсорной плакоды указывают на происхождение основной части (практически всего) слезно-носового протока из предглазничного отрезка инфраорбитального канала органов боковой линии костных рыб, у которых (во всяком случае у кистеперых) с этой частью канала связаны гомологичные lacrymale и septomaxillare кости infraorbitale anterior и rostrale laterale.
Кроме того, нельзя не отметить, что самая проксимальная часть слезно-носового протока образуется дорсальным выступом латерального дивертикула обонятельного мешка, а у H. keyserlingii и вся вентро-латеральная стенка дивертикула, возможно, является продолжением протока, следовательно, этот небольшой участок слезно-носового протока, по-видимому, связан по своему происхождению с органом обоняния.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'