3. Сравнительный обзор развития слезно-носового протока и связанных с ним костей у амфибий
Исследования развития слезно-носового протока у хвостатых и бесхвостых амфибий документируют его чисто кожное происхождение и отсутствие первоначальной связи с органом обоняния и глазом. Судя по литературным данным, совершенно так же обстоит дело и у безногих амфибий (P. Sarasin u. F. Sarasin, 1890). У последних слезно-носовой проток развивается из борозды, проходящей между ноздрей и глазом, которая затем превращается в кожное утолщение. Закладка протока не доходит ни до ноздри, ни до глаза. В дальнейшем эпителиальный валик отшнуровывается от эпидермиса, его передний конец соединяется с якобсоновым органом только во время метаморфоза, а каудальный конец отодвигается от глаза и открывается наружу в особом, присущем безногим амфибиям, щупальце. Следовательно, у всех современных амфибий закладка слезно-носового протока независима в своем возникновении от органов, имеющих с ним дефинитивную связь (от глаза и органа обоняния), и поскольку ее развитие начинается непосредственно в покровном эпителии, первоначальный источник, от которого ведет свое происхождение слезно-носовой проток наземных позвоночных, следует искать среди кожных образований их водных предков.
Закладка слезно-носового протока амфибий возникает в области орбито-назального выступа (flexura lacrymalis) инфраорбитальной сейсмосенсор-ной линии и располагается в непосредственной близости от невромастов этой линии. Развитие закладки протока необычайно сходно с развитием каналов боковой линии костных рыб, хотя и несколько изменено. Но даже у одного и того же вида рыб существуют различия в развитии отдельных каналов боковой линии и связанных с ними костей (Pehrson, 1922, 1940), а также в развитии их у разных видов (Devillers, 1946, 1947, 1948, 1950, 1954, 1965; Lekander, 1949). Обобщенная картина развития каналов боковой линии рыб и относящихся к ним костей выглядит следующим образом. Эпителий будущего канала выдифференцировывается в составе остального эпидермиса и подразделяется на отдельные органы боковой линии - невромасты. Базальная мембрана эпидермиса в местах нахождения невромастов, а также под частями будущего канала, расположенными рядом с невромастами, разрушается. Под разрушенной мембраной скапливается большое количество мезенхимы. Как считает Пирсон (Pehrson, 1940), из эпидермиса в эту мезенхиму мигрируют клетки, из которых в дальнейшем развиваются остеобласты. Девиллер сомневается в подобном происхождении остеобластов, считая, что для выяснения этого вопроса необходимы тщательные экспериментальные исследования и предпочитая пока признавать мезенхиматозное происхождение остеобластов (Devillers, 1948, 1950). Скелетогенная мезенхима либо подстилает покровный эпителий на протяжении одного или нескольких невромастов будущего канала, например, в инфраорбитальной линии, либо представляет собой отдельные парные скопления по бокам каждого невромаста (бинарные закладки Пирсона (Pehrson, 1922)), например, в супраорбиталь-ной линии. Целостность базальной мембраны покровного эпителия восстанавливается, и каждый отдельный невромаст начинает погружаться в глубь тканей, образуя желобок, замыкающийся в канал (рис. 64, А, В). Инвагинировавшие и отшнуровавшиеся от поверхности невромасты (каждый с коротким отрезком канала) соединяются друг с другом в единый канал; сливаются и скопления мезенхимы, окружающие этот канал сплошной мезенхиматозной муфтой, в которой в дальнейшем и возникает костный канал, включающий в себя сейсмосенсорный. Этой части образующейся каналовой кости Девиллер дал название дермо-компонента. От последнего начинает разрастаться собственно тело кости, или мембрано-компонент. Такое подразделение каналовых костей в начале их развития на трубочку и пластинку отмечал ранее и А. Н. Северцов (1925).
Возможен и другой путь образования каналовой кости, наблюдающийся, например, при возникновении frontale у Salmo fario (Devillers, 1947, 1965). При этом под каждым невромастом появляется одинарная простая бластема, связанная с покровным эпителием. Бластемы отдельных невромастов объединяются в общую мезенхимную закладку кости, погружающуюся вглубь и образующую сначала мембрано-компонент (рис. 64, Б). Лишь после возникновения мембрано-компонента происходит инвагинация и отшнурование невромастов, под которыми вторично появляются уже бинарные бластемы, и образование дермо-компонента, т. е. будущего костного канала. В дальнейшем закладки объединяются в единую кость.
Развитие слезно-носового протока амфибий также начинается с дифференцировки части клеток покровного эпителия, а именно клеток его базального слоя и концентрации скелетогенной мезенхимы под закладкой. У бесхвостых амфибий (а, возможно, и у высших хвостатых амфибий: пример - Triton cristatus karelini) произошел временной сдвиг в последовательности этих процессов; у них скопление мезенхимных клеток под местом будущей закладки слезно-носового протока предшествует дифференцировке клеток самой закладки. В отличие от фрагментарности закладки канала боковой линии рыб, обусловленной наличием отдельных органов - невромастов, и возникновения ее одновременно на всем протяжении будущего канала, закладка слезно-носового протока амфибий целостна и сначала появляется на меньшем Отрезке покровного эпителия, постепенно разрастаясь на всю длину протока. Такое изменение хода развития вполне понятно: слезно-носовой проток давно утратил органы боковой линии, обусловливавшие развитие канала из отдельных частей, и перестал быть в результате этого сейсмосенсорным каналом; естественно, что на протяжении длительного филогенетического развития стадия образования его из отдельных отрезков могла выпасть, тем более что уже не было присутствия невромастов, которые могли бы способствовать сохранению этой стадии. Однако у хвостатых амфибий, и не только у наиболее примитивных, но даже у представителей сем. Salamandridae, а также у некоторых бесхвостых амфибий, наблюдается в виде вариаций отдельная самостоятельная закладка добавочной третьей, а иногда и основной - второй, концевой веточки слезно-носового протока, как свидетельство того, что его закладка не всегда была едина.
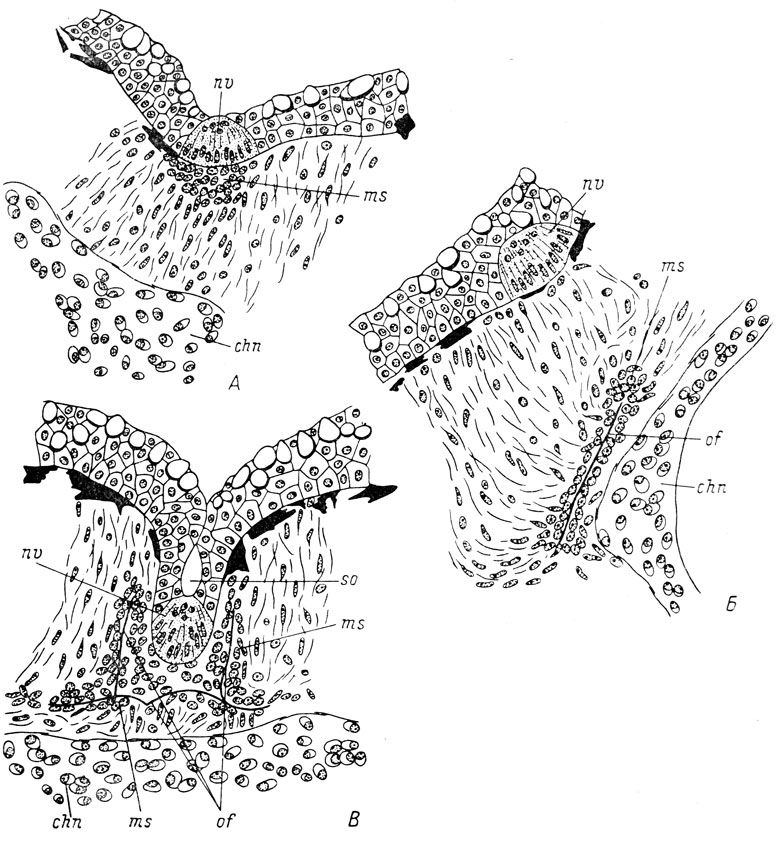
Рис. 64. Развитие каналов боковой линии и связанных с ними костей у Salmo fario. По: Devillers, 1965. А - погружающийся невромаст со скоплением мезенхимы под ним; Б - закладка кости в мезен-химатозной бластеме; В - образование канала и окружение его костью
Несмотря на то что закладка слезно-носового протока во время своего нахождения в покровном эпителии никогда не превращается в канал, а всегда образует плотный внутрикожный тяж, начальные стадии образования этого тяжа (загибание краев клеточного пласта наружу, образование продольной бороздки как предвестника канала), особенно ярко выраженные у низших хвостатых амфибий, весьма напоминают картину образования и инвагинирования канала боковой линии рыб. Незавершение этой картины образованием канала и отшнурованием его опять-таки говорит только о выпадении промежуточных стадий развития и сдвиге возникновения полости на более поздние сроки. Однако у одного из представителей исследованных видов бесхвостых амфибий, причем достаточно примитивного, а именно у Pelobates fuscus, сохранились следы более раннего образования полости канала: у этого вида она появляется (и что чрезвычайно интересно - в виде отдельных, не связанных друг с другом, участков) в то время, когда слезно-носовой проток еще находится на стадии развития внутрикожного тяжа.
Под закладкой слезно-носового протока амфибий точно так же происходит концентрация мезенхимы, как это имеет место под закладкой каналов органов боковой линии рыб. У амфибий не наблюдается нарушения целостности базальной мембраны покровного эпителия под ранней закладкой протока и миграции из нее клеток, превращающихся в дальнейшем в остеобласты, как это отмечено Пирсоном при развитии каналов боковой линии рыб, но при погружении тяжа слезно-носового протока в глубь тканей подстилающая его мезенхима сопровождает проток, окружая его такой же мезенхиматозной муфтой, какой окружены каналы боковой линии. Более того, у самых примитивных хвостатых амфибий в этой мезенхиматозной муфте закладываются и развиваются две кости, охватывающие слезно-носовой проток так же, как окружаются каналы боковой линии рыб дермо-компонентами их каналовых костей. Следовательно, это скелетогенная мезенхима того же типа, что и мезенхимные клетки, окружающие каналы боковой линии рыб, и только пока не установлено происхождение составляющих ее остеобластов, но поскольку оно не выяснено и у рыб, это является в данном случае вопросом второстепенным. Таким образом, у низших хвостатых амфибий две покровные кости lacrymale и septomaxillare, во-первых, развиваются как типичные каналовые кости; во-вторых, они по своему происхождению относятся к той самой инфраорбитальной сейсмосенсорной линии, в области невромастов которой дифференцируется первичная закладка слезно-носового протока. Все эти факты указывают с большой определенностью на происхождение слезно-носового протока амфибий от инфраорбитального канала органов боковой линии рыб.
Безусловно, в восходящих рядах как хвостатых, так и бесхвостых амфибий происходило изменение развития слезно-носового протока как образования, совершенно перестроившегося в связи с полной сменой функции, и исчезала постепенно тесная связь протока с относящимися к нему каналовыми костями. Именно поэтому исследование высших представителей обоих отрядов не может пролить свет на происхождение слезно-носового протока, а плодотворно только изучение наиболее примитивных амфибий и последующее сравнение с высшими, дающее возможность выяснения всего хода эволюционных преобразований.
Сравнительный анализ полученных данных показывает, что отрезок инфраорбитального сейсмосенсорного канала, послуживший источником для возникновения слезно-носового протока наземных позвоночных, исходно не был связан ни с органом обоняния, ни с глазом. Затем возникла первоначальная связь с органом обоняния, а именно с ноздрей, о чем свидетельствуют ход развития слезно-носового протока у низших хвостатых амфибий и всех бесхвостых амфибий, заключающийся в обязательном подрастании переднего конца закладки протока к задне-латеральному краю ноздри, и в лишь затем наступающем последовательном перемещении этого переднего конца по стенке вводного канала к окончательному месту прикрепления - латеральному дивертикулу обонятельного мешка. Хотя у высших хвостатых амфибий вследствие сокращения развития и выпадения некоторых его стадий эта первоначальная связь слезно-носового протока с ноздрей утрачена и его передний конец непосредственно растет к обонятельному мешку, предшествующее существование такой связи доказывается редукцией протока у Pleurodeles waltlii, в ходе которой этот проксимальный (передний) конец совершает в обратном порядке весь путь своего изначального исторического передвижения: от латерального дивертикула обонятельного мешка по стенке вводного канала к ноздре и к покровам. Вполне вероятно, что установление первоначальной связи слезно-носового протока с ноздрей произошло еще на его организационном уровне как канала органов боковой линии у водных предков амфибий, поскольку пример подобного соединения инфраорбитального канала с задней ноздрей имеется у костной ганоидной рыбы Amia calva (Allis, 1889; И. И. Шмальгаузен, 1958д), а Э. И. Воробьевой (1972b) недавно выявлена подобная связь этого же канала с носовой полостью у остеолепиформной кистеперой рыбы Panderichthys rhombolepis. Следовательно, утверждение Ярвика (Jarvik, 1964) о невозможности такой связи противоречит имеющимся фактам.
Хорошо прослеживается в ряду современных амфибий и постепенное исчезновение связи слезно-носового протока с относящимися к нему каналовыми костями, происходившее независимо и параллельно в обоих рассматриваемых отрядах. Если у низших хвостатых амфибий сем. Hynobiidae еще имеются обе каналовые кости lacrymale и septomaxillare, относящиеся к тому отрезку инфраорбитального канала органов боковой линии, из которого филогенетически возник слезно-носовой проток, и развивающиеся в тесной связи с ним, в окружающей его скелетогенной мезенхиме, образуя типичные, хотя и короткие, костные каналы вокруг него, то уже у Ambystomidae и тем более у Salamandridae дело обстоит иначе. Lacrymale, как считается, вообще отсутствует у хвостатых амфибий, исключая Hynobiidae и частично Ambystomidae, а ее место занимается другой покровной костью носо-глазной области черепа - praefrontale, относящейся по своему происхождению не к инфраорбитальному, а к супраорбитальному сейсмосенсорному каналу. У Ambystomidae наличие независимой lacrymale твердо признано только у Dicamptodon, уже относительно Rhyacotriton существуют разногласия (Tihen, 1958; Cloete, 1961, цит. по: Jurgens, 1971; Srinivasachar, 1962), а что касается Ambystoma, то есть только одно указание Папендика (Papendieck, 1954, цит. по: Jurgens, 1971) на вхождение этой кости в состав praefrontale у A. macrodactylum. Однако описанный выше ход развития praefrontale у аксолотля (Ambystoma sp.) подтверждает данные Папендика и показывает, что она может возникать из двух самостоятельных, не связанных друг с другом закладок, причем передне-вентральная из этих закладок образуется в скелетогенной мезенхиме, окружающей среднюю часть слезно-носового протока, в положении, типичном для закладки lacrymale низших хвостатых амфибий. Отсюда можно сделать вывод, что у Ambystoma lacrymale еще не исчезла полностью, а вошла в состав так называемой "praefrontale", сохранив определенную, хотя и не столь тесную, как у примитивных хвостатых амфибий, связь со слезно-носовым протоком, а все семейство Ambystomidae характеризуется наличием слезной кости, либо независимой, либо входящей в состав praefrontale.
Что касается Salamandridae, то судить по данным нормального развития, является ли у них эта кость тоже составной, как у Ambystomidae, и, следовательно, гомологичной одновременно двум покровным костям низших хвостатых амфибий - lacrymale и praefrontale или же истинной praefrontale, трудно. Однако судя по одинаковому развитию этой кости у Ambystomidae и у двух представителей Salamandridae : Triton taeniatus и Pleurodeles waltlii, у которых ее закладка возникает так же, как у аксолотля, на месте типичной lacrymale примитивных хвостатых амфибий, и последующее развитие кости осуществляется тем же самым образом, можно предполагать в согласии с Юргенсом (Jurgens, 1971) ее составную природу и у высших хвостатых амфибий.1 Если учесть, что у всех бесхвостых амфибий lacrymale полностью отсутствует, то получается картина постепенного ослабления связи между слезно-носовым протоком и этой костью, сопровождающегося ее полным исчезновением. Однако это, по-видимому, не совсем так, и придется еще раз вернуться к этому вопросу после анализа развития septomaxillare.
1 (Отклонения в ходе развития "praefrontale" у Triton cristatus karelini, заключающиеся в первоначальной закладке каудальной части кости, а не передне-вентральной, как у других S alamandridae, судя по дефинитивному расположению рассматриваемой кости и ее отношению к слезно-носовому протоку и другим смежным покровным костям, явно недавнего происхождения и не могут ставить под сомнение ее гомологию у всех высших хвостатых амфибий.)
С septomaxillare, на первый взгляд, далеко не такая ясная картина, хотя по данным нормального развития этой кости она бесспорно имеет кожное происхождение как у бесхвостых, так и у хвостатых амфибий, закладываясь в скелетогенной мезенхиме, окружающей проксимальный конец слезно-носового протока у места впадения его в обонятельный мешок и лишь вторично прирастая в некоторых случаях к хрящу обонятельной капсулы. Прежде всего нужно отметить, что septomaxillare в процессе своего исторического развития уходит с поверхности вглубь, переставая быть в прямом смысле покровной костью крыши черепа. Это ее плохо объяснимое перемещение становится понятным, если учесть, что septomaxillare была каналовой костью того отрезка инфраорбитального сейсмосенсорного канала, который превратился в передний (проксимальный) конец слезно-носового протока. При таком истолковании истории кости все становится на свое место. Зависимая в своем возникновении от упомянутого, отрезка инфраорбитального сейсмосенсорного канала (экспериментально доказано, что каналовые кости рыб индуцируются каналами боковой линии (Devillers, 1944, 1946, 1947)) septomaxillare вынуждена была погружаться вглубь вместе с этим отрезком канала, превратившимся в слезно-носовой проток. Это хорошо подтверждается промежуточным (между исходным - у края ноздри и дефинитивным - у латеральной стенки обонятельного мешка) положением закладки этой кости у бесхвостых амфибий, где она возникает в то время, когда передний конец слезно-носового протока еще не закончил своего перемещения и впадает во вводной канал (рис. 62).
В отличие от lacrymale существование septomaxillare у амфибий (она обнаружена у трех семейств хвостатых амфибий: Hynobiidae, Ambystomidae и Plethodontidae, и у всех бесхвостых, у которых она развита значительно сильнее (Lapage, 1928a, 1928b)) обусловлено, по-видимому, не только ее более или менее тесной связью со слезно-носовым протоком, но и приобретением новых функций, среди которых определенную роль играет прикрепление к ней мускулов, сокращающих и расширяющих ноздрю (И. И. Шмальгаузен, 1958г), как об этом упоминалось выше (стр. 97 - 98). Поэтому, пользуясь только данными нормального развития этой кости, нельзя проследить ослабление ее связи со слезно-носовым протоком в филогенетическом аспекте.
Зато данные по нормальному развитию слезно-носового протока у бесхвостых амфибий дают весомые доказательства того, что сильное развитие у последних septomaxillare, вероятно, обусловлено составной природой этой кости, по крайней мере у некоторых Anura, . У бесхвостых амфибий каудальная часть septomaxillare выходит из хрящевой обонятельной капсулы и, продолжаясь на значительное расстояние назад, ограничивает медиально и вентрально среднюю часть слезно-носового протока, т. е. занимает положение lacrymale, которая у Anura, не обнаружена. В связи с таким положением каудальной части septomaxillare очень показательно, что у двух из исследованных видов бесхвостых амфибий, а именно у Реlobates fuscus и Rana temporaria, наблюдалась неоднократно самостоятельная независимая закладка этой каудальной части в положении, типичном для закладки lacrymale - в скелетогенной мезенхиме, окружающей среднюю часть слезно-носового протока.
Поэтому есть все основания полагать, что кость, принимаемая у бесхвостых амфибий за septomaxillare, произошла на самом деле в результате соединения двух покровных костей: lacrymale и septomaxillare, представляющих собой каналовые кости одного и того же инфраорбитального сейсмосенсорного канала. Можно думать, что в этом частичном сохранении находящейся на пути к полному исчезновению кости, совершающемся различными способами у хвостатых и бесхвостых амфибий, играют не последнюю роль остатки формативной зависимости lacrymale 01 деривата инфраорбитального сейсмосенсорного канала, продолжающего свое существование у наземных позвоночных в виде слезно-носового протока.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'