2. Развитие слезно-носового протока у бесхвостых амфибий
Развитие слезно-носового протока у бесхвостых амфибий не имеет столь существенного значения для решения обсуждаемого вопроса о его происхождении, как развитие его у хвостатых амфибий, так как у Anura, , во-первых, сильно видоизменено и усложнено развитие органов боковой линии (И. И. Шмальгаузен, 1955в), что безусловно затрудняет возможность установления происхождения протока от этих органов; во-вторых, у бесхвостых амфибий в настоящее время не вызывает сомнений кожная природа имеющейся у них одной из связанных с протоком костей, а именно septomaxillare (см. стр. 95); в-третьих, у бесхвостых амфибий полностью отсутствует не только вторая, связанная со слезно-носовым протоком кость lacrymale, но и другая покровная кость носо-глазной области черепа, принимаемая у большинства хвостатых амфибий за praefrontale (Stadtmtiller, 1936). Поэтому развитие слезно-носового протока у бесхвостых амфибий будет изложено вкратце, в общей форме, чтобы имелась возможность последующего сравнения.
Эта общая картина развития слезно-носового протока весьма сходна, за исключением деталей, у всех исследованных видов бесхвостых амфибий: Xenopus laevis, Bombina bombina, Pelobates fuscus и Rana temporaria. Все развитие слезно-носового протока захватывает очень небольшой отрезок времени: оно начинается непосредственно перед метаморфозом и в основном развертывается в течение последнего. Самый начальный период развития протока протекает очень быстро и поэтому в большинстве случаев трудно уловим. Он заключается в концентрации мезенхимы под довольно коротким отрезком покровного эпителия, расположенного на небольшом расстоянии от ноздри, сзади и несколько вентральнее нее. Никаких следов дифференциации самого покровного эпителия в месте закладки обнаружить при этом не удается (личинка Pelobates fuscus длиной 51 мм, личинка Rana temporaria длиной 29 мм, рис. 58, А, Б).
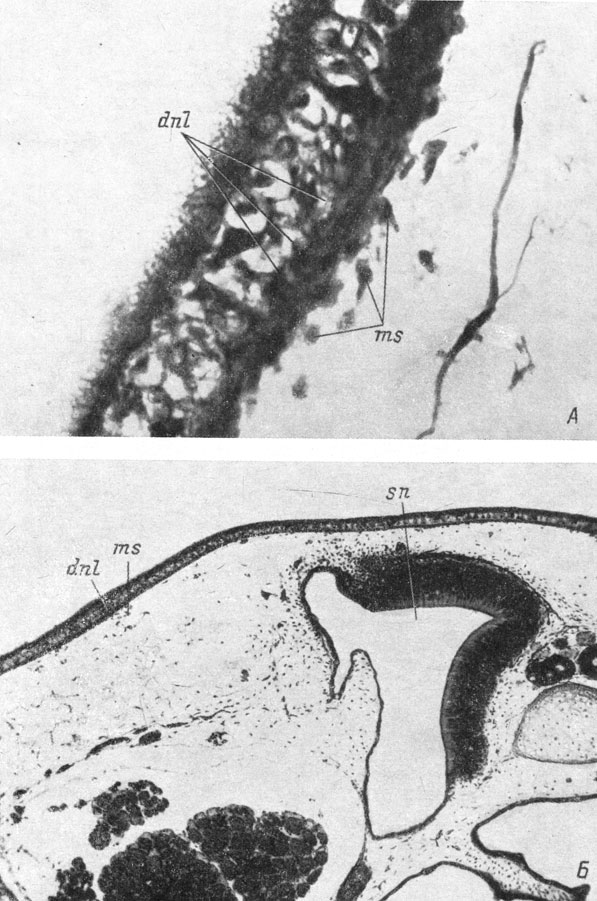
Рис. 58. Ранняя закладка слезно-носового протока у личинки Pelobates fuscus длиной 51 мм (А, увел. 450) и личинки Rana temporaria длиной 29 мм (Б, увел. 66). Поперечные срезы через левую сторону головы
Следующий этап развития слезно-носового протока заключается в концентрации клеток базального слоя покровного эпителия над базальной мембраной, что вызывает незначительное утолщение на внутренней поверхности эпителия (личинка R. temporaria длиной 30 мм, рис. 59) и быстрое распространение закладки вперед до задне-латерального края ноздри. Иногда снаружи, над местом утолщения эпителия, бывает незначительное углубление, но ярко выраженной бороздки, как это имеет место у некоторых хвостатых амфибий, особенно у низших, никогда не образуется. Одновременно с разрастанием закладки вперед происходит и. разрастание ее назад, в поперечном разрезе она в это время представляет собой узкую полоску, расширяющуюся на своем каудальном конце, что можно проследить при просмотре серии срезов.
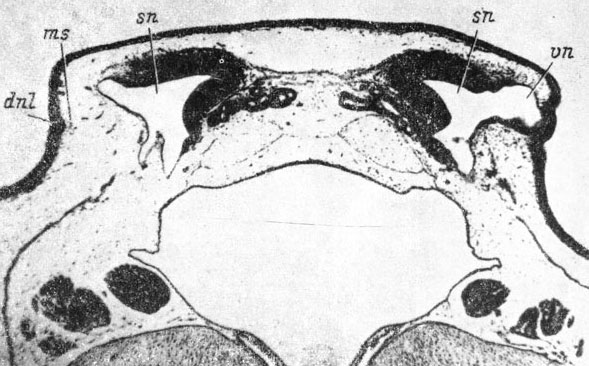
Рис. 59. Поперечный срез через голову личинки Rana temporaria длиной 30 мм. (Увел. 40). Слева - закладка слезно-носового протока
После достижения передним концом закладки слезно-носового протока задне-латерального края ноздри начинается упорядочение клеток базального слоя покровного эпителия, идущее спереди назад, поэтому оно сначала наиболее ярко выражено в районе ноздри (личинка Bombina bombina 42 мм длиной, рис. 60). Это упорядочение клеток сопровождается увеличением утолщения закладки в глубь тканей. Оно приводит к преобразованию переднего, примыкающего к краю ноздри, конца закладки протока в многорядный клеточный пласт, переходящий кзади в плоский оформленный внутрикожный тяж. У личинки Rana temporaria длиной 39 мм этот тяж на каудальном конце немного расширяется и опять переходит в многорядный пласт, на конце раздваивающийся и достигающий уровня заднего края хрящевой обонятельной капсулы. У личинки Pelobates fuscus длиной 44 мм в месте перехода клеточного пласта во внутри-кожный тяж последний, еще проходя внутри кожи, уже начинает отграничиваться от наружных клеток покровного эпителия пограничной мембраной (рис. 61). Продолжая идти внутрикожно, этот тяж уже имеет на поперечном разрезе радиальное расположение клеток, и в некоторых его местах начинается их расхождение, образующее полость. Такое раннее начало образования полости канала изо всех исследованных амфибий наблюдается только у P. fuscus. В своей каудальной половине тяж закладки протока спускается под базальную мембрану покровного эпителия и идет подкожно, прикрепленный к мембране сетью коллагеновых волокон. Каудальный конец тяжа разделяется на две веточки, из которых медиальная кончается в коже, не доходя до уровня переднего края хоаны, латеральная - сзади него. У P. fuscus, а также у Rana temporaria, в некоторых случаях может наблюдаться самостоятельная, не связанная с основным тяжом слезно-носового протока закладка третьей концевой веточки.
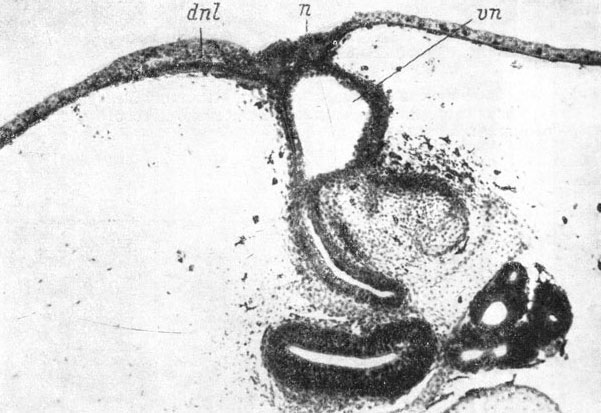
Рис. 60. Поперечный срез через левый орган обоняния личинки Bombina bombina длиной 42 мм. (Увел. 66)
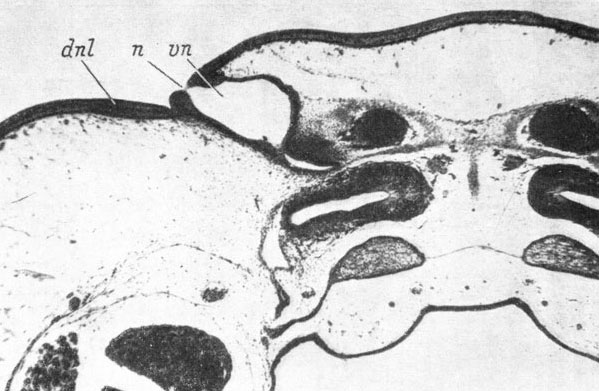
Рис. 61. Поперечный срез через левый орган обоняния личинки Pelobates juscus длиной 44 мм. (Увел. 40)
У личинки Bombina bombina длиной 49 мм при той же степени развития слезно-носового протока его передний конец уже проходит в глубину ноздри внутри ее стенки, а от каудального конца закладки, кзади от окончания в коже обеих веточек, тянется полоса недифференцированного покровного эпителия, отмеченная только скоплением мезенхимы под ней и доходящая до переднего края глаза. На соответствующей описанным стадии 58 у личинки Xenopus laevis слезно-носовой проток представляет собой плотный тяж, отходящий от задне-латерального края ноздри и уже на своем переднем конце спустившийся под покровный эпителий. Базальная мембрана в месте подхода тяжа к коже разрушена. На уровне начала хоаны тяж закладки слезно-носового протока отделяется полностью от покровного эпителия и окруженный собственной мембраной продолжается кзади вплотную под кожей. На уровне заднего края хоаны он немного отходит от покровов в глубь тканей и так идет до своего окончания в коже, достигая уровня заднего края хрящевой обонятельной капсулы. Каудальный конец тяжа слезно-носового протока у X. laevis не раздваивается.
Дальнейшее развитие слезно-носового протока заключается в том, что его передний конец погружается под кожу и начинает соединяться сначала с внутренним краем ноздри (личинка Pelobates fuscus длиной 44 мм), затем этот передний конец передвигается в глубь вводного канала по его латеральной стенке (личинка Xenopus laevis, стадия 59, рис. 62). В это время передний конец слезно-носового протока отделяется как от кожи около края ноздри, так и от стенки ноздри и вводного канала, за исключением самого места прикрепления к ней, и идет от вводного канала через ткани прямо к поверхности тела. Основание протока, соединенное с латеральной стенкой вводного канала, густо окружено скелето-генной мезенхимой, в которой под самым местом впадения протока намечается закладка septomaxillare, расположенная очень далеко от нижнего края хрящевой обонятельной капсулы (личинка Pelobates fuscus длиной 48 мм). И в дальнейшем, по мере разрастания septomaxillare, когда она уже начинает окружать основание слезно-носового протока с трех сторон - дорсально, латерально и вентрально, эта кость все еще находится на достаточном расстоянии от хряща обонятельной капсулы (личинка Xenopus laevis, стадия 59, рис. 62) и ее кожное происхождение не вызывает никаких сомнений. Только позже, когда передний (проксимальный) конец слезно-носового протока, спускаясь по боковой стенке обонятельного мешка, доходит до латерального колена средней носовой полости, разросшаяся septomaxillare срастается дорсально с латеральным краем крыши обонятельной капсулы (личинка Bombina bombina длиной 48 мм, личинка Rana temporaria длиной 31 мм). У Xenopus laevis и Bombina bombina латеральное (или латеро-вертикальное) колено средней носовой полости, от которого дефинитивно отходит своим проксимальным концом слезно-носовой проток, образует дополнительное передне-боковое объемистое расширение. Последнее с равной степенью вероятности можно считать и расширенным проксимальным концом самого протока.
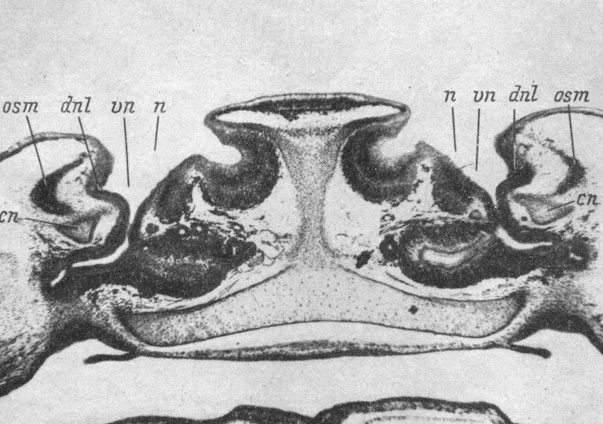
Рис. 62. Поперечный срез через голову личинки Xenopus laevis длиной 55 мм, стадия 59, на уровне впадения переднего конца слезно-носового протока во вводной канал органа обоняния. (Увел. 40)
После того как передний конец слезно-носового протока достигает края ноздри и стенки вводного канала, он быстро начинает погружаться в ткани на всем своем протяжении, оставаясь в то же время связанным с базальной мембраной покровного эпителия коллагеновыми волокнами, и одновременно приобретает полость, образование которой начинается от полости обонятельного мешка, а диетальный конец протока вместе с ветвями канализируется в последнюю очередь. У всех исследованных бесхвостых амфибий обе ветви дистального конца протока открываются у основания нижнего века наружу: медиальная веточка кончается у угла глаза, а латеральная проходит далее каудально, примерно до середины глаза. Исключение составляет X. laevis, у этого вида диетальный конец слезно-носового протока не разветвляется и открывается наружу одним отверстием, помещающимся на конце особого сосочка, или щупальца, находящегося под глазом. Развитие слезно-носового протока заканчивается к концу метаморфоза.
Остается сказать еще несколько слов о дефинитивном состоянии и положении septomaxillare. Эта кость у бесхвостых амфибий хотя и несколько варьирует в своем строении у разных видов, но в целом значительно сильнее развита, чем у низших хвостатых амфибий, у которых она также имеется. Как правило, в этой кости можно различить три части (Maas, 1945). Ее передне-дорсальный или же дорсальный край может поддерживать вентральную стенку ноздри и вводного канала, подходя очень близко к поверхности тела (Bombina bombina), либо - только стенку вводного канала (Rana temporaria), либо просто представлять собой восходящую пластинку, лежащую латерально от боковой стенки обонятельного мешка и не достигающую вводного канала (Xenopus laevis, Pelobates fuscus). Далее кзади располагается средняя часть septomaxillare, которая окружает не только проксимальный конец слезно-носового протока, но и латеро-вертикальное колено средней носовой полости (infundibulum), от которого он отходит. Эта часть septomaxillare почти целиком находится внутри хрящевой обонятельной капсулы и часто дает еще длинный отросток, охватывающий латеро-вертикальное колено средней полости с передней стороны и заходящий медиально от него. Этот отросток septomaxillare можно рассматривать как pr. dermintermedius остеолепиформных кистеперых рыб. Средняя часть обычно прирастает своими дорсальной и вентральной сторонами к хрящу обонятельной капсулы, но в то же время между костью и хрящом может сохраняться прослойка соединительной ткани.
Латеро-каудальная часть septomaxillare сопровождает слезно-носовой проток на довольно значительное расстояние после его выхода из обонятельной капсулы, и поэтому эта третья часть кости лежит много более поверхностно, чем другие части (у Bombina bombina эта латеро-каудальная часть отсутствует, и проток окружен septomaxillare только в своей расширенной проксимальной части, находящейся внутри обонятельной капсулы). Сопровождая слезно-носовой проток по выходе из капсулы латеро-каудальная часть septomaxillare окружает хотя и не полностью, а только медиально и вентрально его среднюю часть, т. е. выполняет функцию lacrymale низших хвостатых амфибий. Восходящий отросток maxillare, прикрывающий латерально и вентрально среднюю часть слезно-носового протока у высших хвостатых амфибий, у бесхвостых не дорастает до протока.
В связи с описанным строением и прохождением латеро-каудальной части septomaxillare очень интересен тот факт, что в некоторых случаях у Pelobates fuscus и Rana temporaria отмечается независимая закладка этой части кости в скелетогенной мезенхиме, окружающей среднюю часть слезно-носового протока. Во всех этих случаях закладка кости располагалась медиально и немного вентрально от слезно-носового протока, т. е. очень сходно с закладкой lacrymale у низших хвостатых амфибий. В одном случае у R. temporaria две независимые кости еще сохранялись у уже закончившей метаморфоз сеголетки. При этом кость, находившаяся с медиальной и вентральной стороны средней части слезно-носового протока, не имела никакой связи с основной частью septomaxillare и располагалась как типичная lacrymale низших хвостатых амфибий (рис. 63 ср. с рис. 46, стр. 97). В то же время в норме у бесхвостых амфибий на этом месте обычно размещается латеро-каудальный конец septomaxillare. Такие варианты развития кости указывают на составное происхождение septomaxillare у Anura.
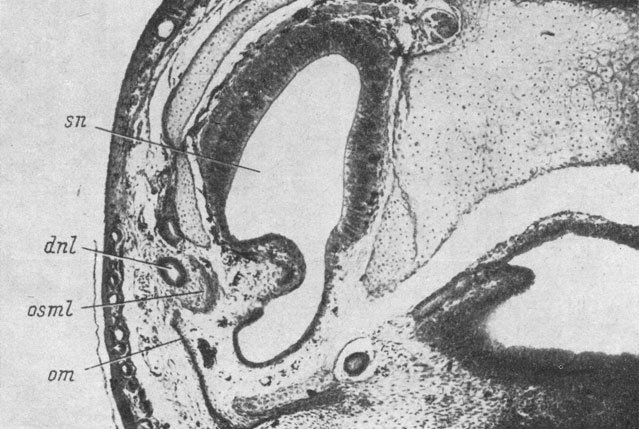
Рис. 63. Поперечный срез через левый орган обоняния сеголетки Rana temporaria длиной 13 мм. (Увел. 66)
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'