1. Развитие слезно-носового протока у хвостатых амфибий
В своей работе по исследованию развития слезно-носового протока у хвостатых амфибий И. И. Шмальгаузен (1958г) привел с исчерпывающей полнотой описание развития этого образования у Hynobius keyserlingii, развитие протока у других видов дано фрагментарно. Поэтому в настоящем изложении по H. keyserlingii приведены основные данные И. И. Шмадьгаузена с небольшими дополнениями, описание развития слезно-носового протока у других видов - оригинальное.
Hynobiidae. У H. keyserlingii самая ранняя морфологически различимая закладка слезно-носового протока распознается в покровном эпителии у личинок 14 мм длиной в области орбито-назального выступа (flexura lacrymalis) инфраорбитальной плакоды органов боковой линии. Эту закладку можно различить по некоторому сгущению клеток покровного эпителия и скоплению под ней мезенхимных клеток. У личинок 16 мм длиной (рис. 45) закладка протока уже образует небольшое утолщение в глубь тканей, а клетки базального слоя эпителия обладают более высокой цилиндрической формой. Одновременно закладка слезно-носового протока вытягивается в направлении ноздри и в направлении глаза, превращаясь во внутрикожный пласт. На всем своем протяжении этот пласт подстилается скоплением мезенхимных клеток.
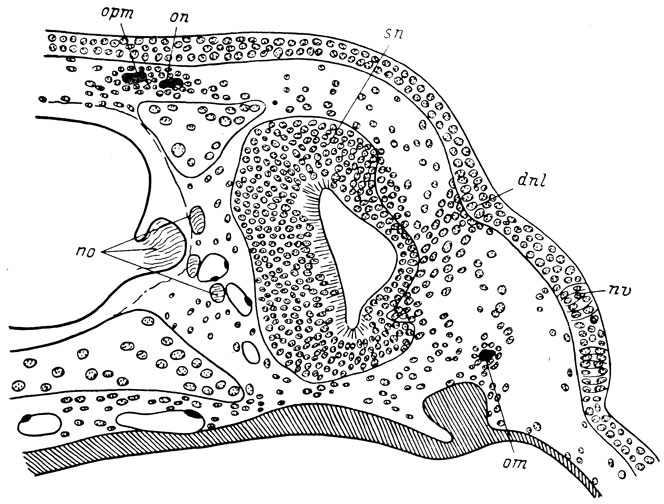
Рис. 45. Закладка слезно-носового протока у личинки Hynobius keyserlingii длиной 16 мм. По: И. И. Шмальгаузен, 1958 г.
У личинок длиной 18 - 20 мм закладка слезно-носового протока начинает компановаться в более плотный внутрикожный тяж, который к заднему (глазному) концу расширяется, постепенно теряя ясные границы и переходя в утолщение век. Впереди она доходит до наружного края ноздри. У личинок длиной 20 - 24 мм передний конец закладки протока продолжается по задней стенке вводного канала (преддверия) и достигает латерального дивертикула обонятельного мешка. Почти одновременно с этим начинается погружение плотного внутрикожного тяжа закладки слезно-носового протока в глубь тканей и отделение его от вводного канала ноздри и от покровного эпителия. Подстилающая закладку протока, как и весь покровный эпителий, базальная мембрана погружается вместе с тяжом; часть ее, окружающая тяж слезно-носового протока, отрывается от основной мембраны, давая тем самым собственную мембрану протока. Вместе с тяжом погружаются и мезенхимные клетки, образующие интенсивное скопление вокруг него на всем его протяжении.
У личинок длиной примерно 28 - 30 мм начинается образование полости слезно-носового протока, причем процесс этот идет спереди от обонятельного мешка по направлению к дистальному расширенному концу закладки протока, который в это время дифференцируется на две отдельные ветви, оканчивающиеся в коже. В них тоже образуется полость, а всего позднее, примерно к началу метаморфоза, канализируется средняя часть слезно-носового протока. Формируясь окончательно к концу метаморфоза, слезно-носовой проток открывается наружу у края век более короткой (латеральной) и более длинной (медио-дорсальной) ветвями. Довольно часто латеральная ветвь может отсутствовать.
Закладки lacrymale и septomaxillare возникают в мезенхимной муфте, окружающей слезно-носовой проток, практически одновременно у личинок длиной 26 мм. Закладка lacrymale располагается в средней части протока (рис. 46) сначала вентро-медиально от него, а затем разрастается вокруг протока, охватывая его в виде кольца, а позже (во время метаморфоза) в виде трубочки. Дистальное разветвление слезно-носового протока лежит на задней части дорсальной поверхности lacrymale.
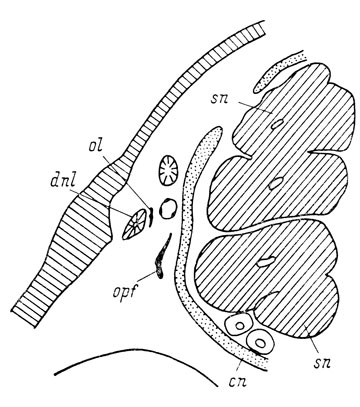
Рис. 46. Закладка lacrymale у личинки Hynobius keyserlingii длиной 26 мм. По: И. И. Шмалъгаузен, 1958 г.
Закладка septomaxillare образуется у вентро-латеральной стенки переднего конца слезно-носового протока в месте его впадения в латеральный дивертикул обонятельного мешка, но на достаточном расстоянии от хряща обонятельной капсулы и совершенно независимо от него (рис. 47). Она возникает как типичное мембранное окостенение в скоплении мезенхимы, окружающей проток. Эта косточка, быстро разрастаясь, окружает проксимальный конец слезно-носового протока полным кольцом и остается независимой от хряща обонятельной капсулы вплоть до метаморфоза, во время которого ее вентральный край прирастает к вентро-каудальному краю fenestra narina. Это вторичное прирастание septomaxillare к хрящу И. И. Шмальгаузен объясняет функциональными требованиями: musculus constrictor naris и m. dilatator naris прикрепляются не только к краям fenestra narina, но и к septomaxillare, и их функционирование возможно только при условии прочного фиксирования этой кости на хряще неподвижной обонятельной капсулы.
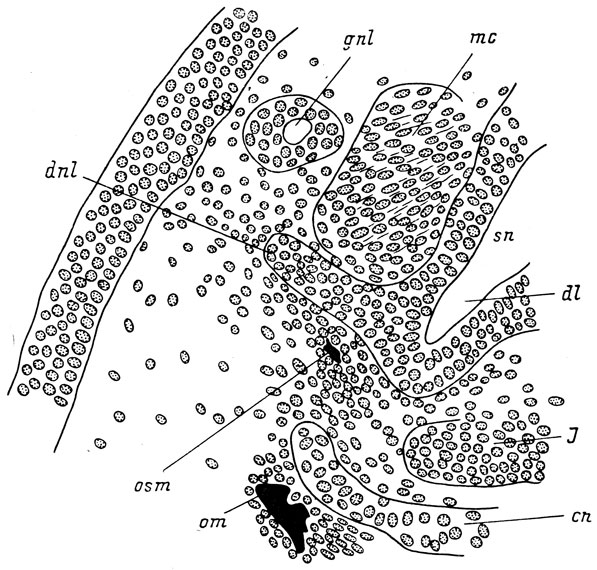
Рис. 47. Закладка septomaxillare у личинки Hyriobius keyserlingii длиной 28 мм. По: И. И. Шмальгаузен, 1958 г.
У Ranodon sibiricus закладка слезно-носового протока также располагается в области орбито-назального выступа инфраорбитальной плакоды органов боковой линии, и первые морфологические признаки ее заключаются в упорядочении одного ряда клеток базального слоя покровного эпителия и концентрации мезенхимы непосредственно под закладкой (личинки длиной 24 - 28.5 мм). Нужно отметить, как указывал на это и И. И. Шмальгаузен (1958 г), что развитие слезно-носового протока и связанных с ним костей идет очень неравномерно: зачастую более крупные личинки имеют менее дифференцированный проток, может наблюдаться разница в этом отношении также между правой и левой сторонами одного экземпляра.
У личинок длиной 28.5 - 29.8 мм сохраняющая такой же вид закладка протока становится больше в своем протяжении и увеличивается количество мезенхимы под ней. У личинок длиной 31 - 34.5 мм клетки базального слоя закладки начинают перегруппировываться во внутрикожный многорядный пласт, края которого изгибаются к поверхности, так что снаружи образуется углубление, а изнутри - выпячивание закладки в подлежащую ткань. Углубление хорошо просматривается при наружном осмотре в виде продольной бороздки, обозначающей прохождение закладки слезно-носового протока. Эта картина создает полное впечатление готовящегося впячивания и отшнурования закладки протока с превращением срединной борозды в его полость. Однако никакого отшнурования не происходит, а закладка слезно-носового протока, разрастаясь сначала назад, по направлению к глазу, а затем уже вперед, по направлению к ноздре, преобразуется в средней части своего протяжения во вполне оформленный, круглый на поперечном разрезе, плотный внутри-кожный тяж. Кпереди этот тяж переходит в такой же многорядный эпителиальный пласт, которым была представлена закладка протока у более молодых личинок, немного не достигающий ноздри (личинки длиной 35 - 37 мм); сзади он также расширяется в пласт, в отдельных случаях доходящий до уровня переднего края хоаны. У личинок длиной 38 мм весь передний отрезок закладки слезно-носового протока преобразуется во внутрикожный тяж, который доходит до заднего края ноздри.
Достигнув заднего края ноздри, тяж слезно-носового протока отделяется от кожи и передний конец его начинает спускаться вглубь по задней стенке ноздри (личинки длиной 39 - 44 мм). В месте погружения тяжа базальная мембрана прерывается и быстро срастается вновь, а часть ее, первоначально подстилавшая тяж, образует собственную мембрану слезно-носового протока, замыкаясь вокруг него. Снаружи пограничной мембраны протока располагается толстый слой скелетогенной мезенхимы. Средняя часть тяжа тоже спускается под кожу, но продолжает оставаться соединенной с ней благодаря погрузившейся базальноймембране, на которой она как бы подвешена. Каудальная часть закладки протока, также превратившаяся в тяж, еще остается внутрикожной и на своем заднем конце раздваивается, причем латеральная часть раздвоенного конца заканчивается раньше медиальной, которая доходит до уровня переднего края хоаны.
У личинки длиной 48 мм передний конец плотного тяжа слезно-носового протока, клетки которого ориентированы на поперечном срезе радиально, впадает в вентро-латеральную стенку вводного канала ноздри довольно далеко от ее края и продолжается в толще стенки до латеральной стенки главной носовой полости. Средняя часть тяжа протока проходит довольно глубоко под кожей, но на всем своем протяжении соединена с базальной мембраной покровного эпителия коллагеновыми волокнами и особенно плотно окружена скелетогенной мезенхимой. В этой мезенхиме, у места перехода средней части слезно-носового протока в заднюю, непосредственно перед раздвоением тяжа, медиально от протока намечается закладка lacrymale. Каудальная часть протока состоит из двух ветвей, уже спустившихся под кожу, из которых медиальная, более длинная, веточка заканчивается на уровне передней части хоаны.
У личинки длиной 51.5 мм передний конец плотного слезно-носового протока уже впадает в латеральный дивертикул (боковой носовой желобок) обонятельного мешка. Место впадения окружено густым скоплением скелетогенной мезенхимы. Закладка lacrymale разрослась вперед от разветвления протока вдоль его медиальной стенки и заходит также под него, на вентральную сторону. В начале метаморфоза (личинка длиной 60 мм) lacrymale разрастается еще дальше вперед, подстилая слезно-носовой проток снизу, а в своей задней части, перед разветвлением протока, охватывает его также с медиальной и латеральной сторон. Несколько позже, в начале среднего периода метаморфоза (личинка длиной 66 мм), в скелетогенной мезенхиме, подстилающей проксимальный (передний) конец протока у места его впадения в латеральный дивертикул обонятельного мешка, начинает дифференцироваться закладка septo-maxillare. Эта закладка, как и у Hynobius keyserlingii (рис. 47), находится на очень большом расстоянии от нижнего края fenestra narina хрящевой обонятельной капсулы, кроме того, между этим краем и закладкой septomaxillare, которая возникает гораздо позже, чем у Н. keyserlingii, у Ranodon sibiricus уже находится закладка m. dilatator naris (рис. 68, Б). Lacrymale в это время разрастается с латеральной стороны слезно-носового протока и в своей передней части. В этот же период происходит быстрая канализация протока, причем образование полости может начинаться с его проксимального конца, а может, как отмечал И. И. Шмальгаузен (1958 г), происходить и с дистального конца.
К середине метаморфоза (личинка длиной 74 мм) septomaxillare охватывает проксимальный конец слезно-носового протока снизу, сверху, а по мере его отхождения от латерального дивертикула также и с медиальной стороны, образуя почти полное кольцо вокруг основания протока и находясь еще на достаточно большом расстоянии от хряща обонятельной капсулы. Lacrymale в это время также охватывает проток почти полным кольцом, разрастаясь над ним. Во второй половине метаморфоза (личинка длиной 78 мм) septomaxillare окружает проксимальный конец слезно-носового протока полным кольцом и прирастает своей вентральной стороной к нижнему краю fenestra narina хрящевой обонятельной капсулы. Lacrymale охватывает среднюю часть протока также полным кольцом, образуя короткую трубочку. У взрослых экземпляров слезно-носовой проток открывается медиальной ветвью у края верхнего века с его внутренней стороны, латеральная ветвь немного не доходит до края века и открывается в коже вблизи глаза. Нужно отметить, что разветвление дистального конца слезно-носового протока наблюдается не у всех экземпляров, латеральная веточка может отсутствовать, а может и закладываться самостоятельно, безо всякой связи с основным стволом протока. Точно так же и septomaxillare не всегда образует полное кольцо вокруг основания протока, на что указывал и И. И. Шмальгаузен (1958 г).
У третьего представителя сем. Hynobiidae - Onychodactylus fischeri - не известны ни сроки, ни места откладки икры. В связи с этим исследование развития слезно-носового протока проводилось на отловленных в природе личинках, величина которых очень варьирует и не всегда больший размер личинки соответствует большей степени дифференцировки протока.
Первая морфологически распознаваемая закладка слезно-носового протока отмечена у личинки длиной 34 мм. Место закладки маркируется небольшим подкожным скоплением мезенхимы и представляет собой полоску еще недифференцированного покровного эпителия, располагающуюся как раз над якобсоновым органом и далеко не доходящую как до ноздри, так и до глаза. У личинки длиной 37 мм эта полоска уже состоит из упорядоченного внутреннего слоя клеток, под которым наблюдается значительное скопление мезенхимы. Затем по всей длине закладки, по ее средней линии, возникает продольная, ярко выраженная, наружная бороздка (личинки длиной 41 и 44 мм), а сам клеточный пласт выпячивается под кожу, так что создается полное впечатление готовящегося отшнурования (рис. 48). Однако последнего не происходит, а клеточный пласт, начиная со своего переднего конца, преобразуется во внутрикожный плоский тяж, в то время как наружная бороздка сглаживается (личинка длиной 42 мм). Далее наступает разрастание закладки протока вперед и назад, а точнее, постепенная дифференциация клеток по той же схеме: упорядоченный клеточный пласт, образование наружной бороздки, превращение во внутрикожный тяж. У личинки длиной 50 мм передний конец закладки слезно-носового протока в виде упорядоченного клеточного пласта, подостланного мезенхимным скоплением, лишь немного не доходит до заднего края ноздри. Кзади этот пласт переходит во внутрикожный плоский тяж, вдающийся под кожу и на своем заднем конце раздваивающийся, причем медиальная часть представляет собой внутрикожный пласт, а латеральная - уже подкожный, подмембранный округлый тяж, заканчивающийся в коже.
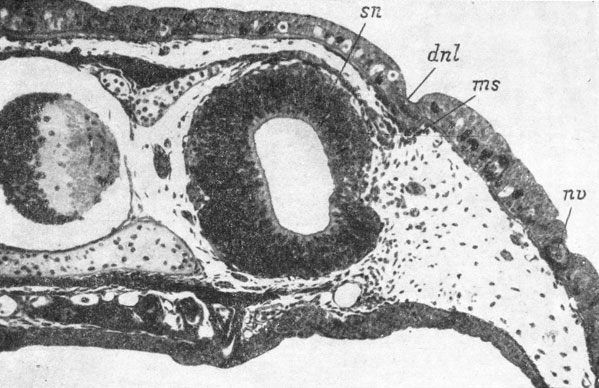
Рис. 48. Закладка слезно-носового протока у личинки Onychodactylus fischeri длиной 41 мм. (Увел. 66)
Затем передний конец закладки протока, все еще имеющий вид клеточного пласта, дорастает до заднего края ноздри, кзади он переходит в плоский тяж, который постепенно превращается в округлый и спускается под кожу, а далее и под базальную мембрану, но пока лежит вплотную под ней. Еще дальше кзади тяж погружается глубже и отделяется от мембраны мезенхимными клетками, но продолжает быть связанным с ней коллагеновыми волокнами. Каудальный конец тяжа разделяется на две веточки, из которых медиальная заканчивается в коже чуть впереди латеральной (личинка длиной 47 мм).
Передний конец закладки слезно-носового протока преобразуется из пласта в тяж и перемещается внутрь по задней стенке вводного канала, впадая в него недалеко от края ноздри. Одновременно вся закладка погружается в глубь тканей и отделяется от кожи мезенхимой на всем своем протяжении (личинка длиной 45 мм). Теперь закладка слезно-носового протока представляет собой лежащий в тканях округлый плотный тяж, в котором начинают образовываться отдельные, пока не связанные друг с другом полости. Передний конец постепенно перемещается вглубь по задней стенке вводного канала к главной полости обонятельного мешка (личинки длиной 52, 53, 55 мм). Когда передний конец протока начинает присоединяться к обонятельному мешку в месте перехода вводного канала в главную полость, в густом скоплении мезенхимы, окружающем заднюю половину тяжа, с медио-вентральной стороны от него и перед разветвлением появляется закладка lacrymale (личинка длиной 52 мм).
Затем передний конец слезно-носового протока перемещается к месту своего дефинитивного впадения в обонятельный мешок - латеральному дивертикулу главной полости и непосредственно под местом соединения с последним, в толще мезенхимы, на очень большом расстоянии от хряща обонятельной капсулы закладывается septomaxillare (рис. 49). Одновременно проток глубоко погружается в ткани на всем своем протяжении. Lacrymale разрастается и окружает заднюю половину протока и место его разветвления с латеральной, вентральной и медиальной сторон. Единой полости протока еще нет (личинки длиной 56 и 65 мм).
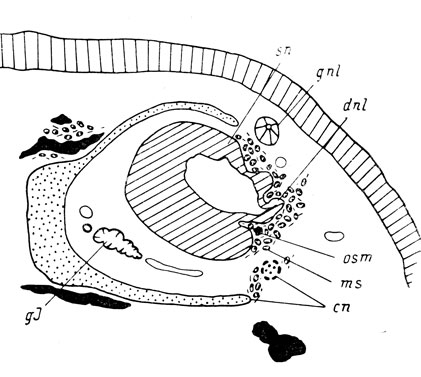
Рис. 49. Закладка septomaxillare у личинки Onychodactylus jischeri длиной 56 мм. (Увел. 66)
У личинки длиной 70 мм septomaxillare полукольцом охватывает основание слезно-носового протока спереди, снизу и сзади и дает впереди длинный изогнутый отросток pr. dermintermedius, заходящий под латеральный дивертикул обонятельного мешка в глубь обонятельной капсулы. Lacrymale образует почти полную трубочку вокруг задней половины протока, открытую только с наружно-верхнего угла. Слезно-носовой проток канализован на всем своем протяжении.
Более старшие личинки, находящиеся в периоде метаморфоза, описаны И. И. Шмальгаузеном (1958 г). Личинка длиной 75 мм (начало метаморфоза) сходна по степени развития слезно-носового протока с вышеописанной личинкой длиной 70 мм. У личинки 85 мм длиной (конец метаморфоза) lacrymale охватывает проток в виде трубки на довольно большом протяжении, а septomaxillare представляет собой наиболее полное развитие этой кости у хвостатых амфибий (рис. 50), так как имеет типичный для нее длинный отросток, вдающийся в обонятельный мешок между его главной полостью и латеральным дивертикулом, названный Вегнером (Wegner, 1922) pr. intrafenestralis narialis у бесхвостых амфибий и у рептилий и Ярвиком - pr. dermintermedius покровной кости rostrale laterale у Eusthenopteron, гомологизируемой им с septomaxillare Anura, и Amniota (Jarvik, 1942). Таким образом, O. fischeri демонстрирует единственный среди хвостатых амфибий пример полного развития septomaxillare вместе с pr. dermintermedius.
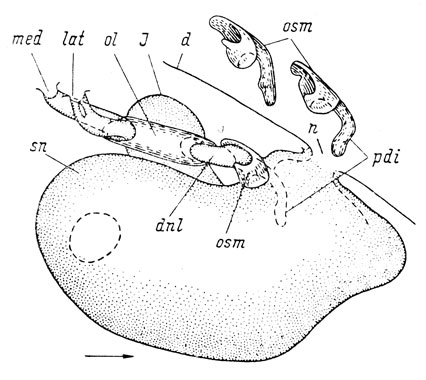
Рис. 50. Реконструкция левого обонятельного мешка личинки Onychodactylus fischeri длиной 85 мм (конец метаморфоза). Вид с дорсальной стороны. Вверху показаны отдельно septomaxillare обеих сторон, правая кость дана в зеркальном отображении. Стрелка указывает передний конец морды. По: И. И. Шмальгаузен, 1958 г.
Ambystomidae. Первая морфологически различимая закладка слезно-носового протока аксолотля (Ambystoma sp.) появляется у личинок длиной 25 - 28 мм (стадии 51 - 52 по Гаррисону (Воронцова и др., 1952)) в виде упорядочения базального слоя клеток покровного эпителия. Она располагается несколько каудальнее, чем у Hynobiidae, над якобсоновым органом обонятельного мешка, и вначале имеет небольшое протяжение. С момента появления закладки протока под ней наблюдается скопление мезенхимных клеток. На стадиях 52 - 56 (личинки длиной 28 - 32 мм) закладка разрастается назад, оставаясь в своей передней половине однорядной и более широкой, а в каудальной половине начинает перестраиваться в многорядный пласт, что сопровождается углублением ее в подлежащую ткань и образованием выпячивания на внутренней стороне покровного эпителия (рис. 51). Углубление на его наружной стороне в месте образования закладки протока возникает не сразу, а к стадии 55. На стадии 57 (личинка длиной 39 мм) передний конец закладки слезно-носового протока в виде упорядоченного ряда клеток базального слоя покровного эпителия располагается над местом отхождения якобсонова органа от обонятельного мешка. Далее кзади она преобразуется в округлый тяж, сильно впячивающийся под кожу, в каудальной же части переходит в более широкий и плоский многорядный пласт, конец которого почти доходит до уровня переднего края хоаны.
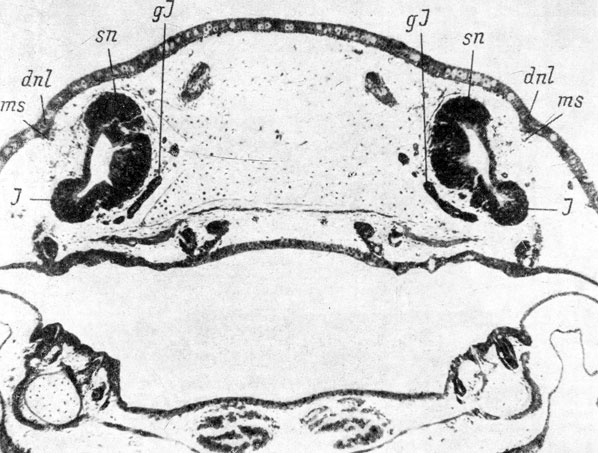
Рис. 51. Закладка слезно-носового протока у аксолотля длиной 32 мм, стадия 55. (Увел. 40)
На стадии 58 (личинки длиной 40 - 41 мм) закладка протока на всем своем протяжении имеет вид округлого подкожного тяжа, передний конец которого, оставаясь на уровне места отхождения якобсонова органа от обонятельного мешка, начинает расти вглубь, разрывая базальную мембрану (рис. 52). Каудальный конец тяжа раздваивается. На стадии 60 (личинки длиной 39 - 41.5 мм) вся средняя часть закладки слезно-носового протока лежит уже под кожей в виде тяжа, отделенного от нее собственной мембраной. Передний конец тяжа продолжает расти в направлении обонятельного мешка. На той же стадии 60 (личинка длиной 42.5 мм) передний, теперь уже собственно проксимальный, конец тяжа срастается с латеральным дивертикулом обонятельного мешка, но еще между тем и другим видны ясные границы. Средняя часть закладки протока отщепилась от покровного эпителия и погрузилась в ткани, и только каудальная часть еще остается в виде подкожного тяжа. Погрузившийся отрезок протока обладает собственной базальной мембраной и окружен со всех сторон мезенхимой. На этой же стадии развития, но у еще более крупных личинок (длина 51.9 мм) в скелетогенной мезенхиме, окружающей каудальный отрезок средней части слезно-носового протока и его заднюю, также отделившуюся от кожи часть, в месте разветвления возникает закладка косточки, находящаяся медиально от протока (рис. 53) совершенно так же, как располагается закладка lacrymale у Hynobiidae (рис. 46), и проходящая между протоком и хрящом обонятельной капсулы. Однако эта кость не обрастает слезно-носовой проток, как это имеет место у низших хвостатых амфибий, а, оставаясь в месте закладки проходящей медиально от протока пластинкой, кроме того, разрастается кзади и дорсально, вдоль хряща обонятельной капсулы и, загибаясь дорсо-медиально, налегает своим дорсальным краем на покрывающую крышу обонятельной капсулы frontale (стадия 61, личинка длиной 62 мм).
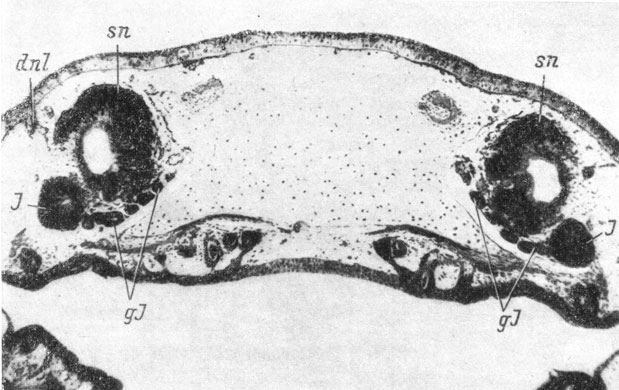
Рис. 52. Поперечный срез головы аксолотля длиной 41 мм, стадия 58, на уровне переднего конца слезно-носового протока. (Увел. 40)
Таким образом, у аксолотля кость, закладывающаяся, как типичная lacrymale низших хвостатых амфибий, разрастаясь, занимает своей дорсо-каудальной частью положение их другой покровной кости, а именно praefrontale (см. рис. 67, А), в качестве каковой она обычно и рассматривается у более высоко организованных хвостатых амфибий. У личинки длиной 65 мм, находящейся на той же стадии развития 61, на одной стороне обсуждаемая кость состоит из двух частей, соответствующих по положению lacrymale и praefrontale низших хвостатых амфибий и соединенных между собой тонкой костной перемычкой, а на другой стороне она вообще представляет собой две отдельные костные закладки, находящиеся в типичных для закладок lacrymale и praefrontale положениях и связанные между собой только скелетогенной мезенхимой (рис. 54). Все развитие этой кости у аксолотля, принимаемой у большинства хвостатых амфибий за praefrontale, показывает, что она имеет составную природу и включает в себя как lacrymale, так и praefrontale низших хвостатых амфибий.
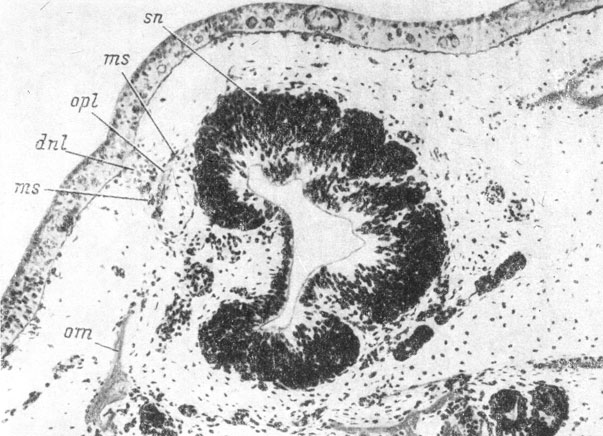
Рис. 53. Поперечный срез левой стороны головы аксолотля длиной 51.9 мм, стадия 60, на уровне закладки praefronto-lacrymale. (Увел. 66)
Старшей из исследованных была личинка длиной 76 мм, соответствовавшая той же стадии развития 61. У нее не было еще никаких признаков образования septomaxillare, кроме плотного окружения проксимального конца слезно-носового протока скелетогенной мезенхимой. Хотя, по литературным данным, эта кость у аксолотля имеется (Bruner, 1902; Noble, 1921; Stadtmuller, 1936), она, вероятно, образуется у более взрослых особей. Слезно-носовой проток также оставался еще плотным тяжом и не обнаруживал признака образования полости, а его разветвленный дистальный конец доходил до уровня передней трети хоаны, причем обе веточки были одинаковой длины и заканчивались в коже друг над другом.
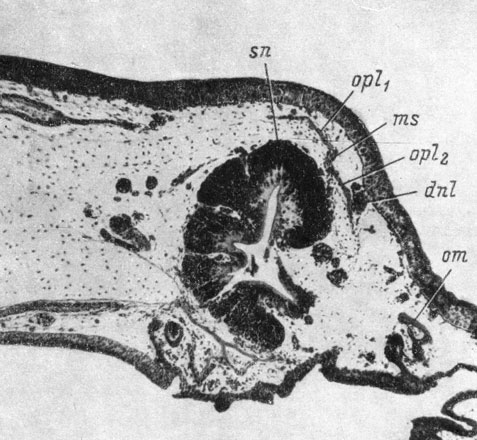
Рис. 54. Поперечный срез правой стороны головы аксолотля длиной 65 мм, стадия 61, на уровне двойной закладки praefronto-lacrymale. (Увел. 40)
Salamcindridae. Развитие слезно-носового протока у Triton taeniatus характерно тем, что его первые стадии сокращены и протекают очень быстро. На стадии 58 по Глюкзон (личинки длиной 15 - 16 мм) еще нет никаких следов закладки протока. На стадии 59 (личинки длиной 15 - 16.5 мм) закладка слезно-носового протока уже имеет вид хорошо оформленного, круглого в поперечном сечении, эпителиального тяжа, который почти на всем своем протяжении наполовину вдается в глубь тканей, выгибая внутрь базальную мембрану покровного эпителия, и только на своем каудальном конце, где он подразделяется на две веточки, эти два тяжика проходят внутрикожно. По всей длине тяжа слезно-носового протока, расположенного как раз над якобсоновым органом, по наружной поверхности покровного эпителия проходит бороздка, более глубокая в задней половине закладки. Под закладкой протока наблюдается скопление мезенхимных клеток.
На стадиях 60 - 61 (личинки длиной 22 - 24 мм) передний конец тяжа протока, не доходя до ноздри, спускается вглубь, подрастает к обонятельному мешку и срастается непосредственно с латеральной стенкой его главной полости, так как выпячивание латерального дивертикула в это время еще отсутствует. Средняя и задняя части слезно-носового протока на этих стадиях развития представляют собой подкожный тяж, плотно прилегающий к базальной мембране покровного эпителия, и только в месте раздвоения тяжа на веточки базальная мембрана прерывается, и они оканчиваются в коже.
В предметаморфозный период (личинки длиной 26 - 34.5 мм) слезно-носовой проток на всем своем протяжении (кроме концевых веточек) отделяется от кожи. Проксимальный (передний) его конец отходит от образовавшегося латерального дивертикула обонятельного мешка. У некоторых особей в этом периоде начинается образование полости протока в направлении от переднего конца к заднему, при этом канализируются его проксимальная и средняя части, у других этот процесс задерживается до середины метаморфоза. В скелетогенной мезенхиме, окружающей каудальный конец средней части протока, перед его разветвлением, дорсо-медиально от протока и рядом с хрящом обонятельной капсулы, образуется закладка косточки очень сходно с тем, что наблюдается у аксолотля. В начале метаморфоза (личинка длиной 37 мм) эта кость разрастается дорсально и каудально вдоль хряща обонятельной капсулы и почти доходит до латерального края frontale, покрывающей крышу обонятельной капсулы.
В конце первой половины метаморфоза (личинка длиной 38 мм) происходит дорсальное разрастание восходящего отростка maxillare, который окружает среднюю часть слезно-носового протока с его латеральной и вентральной сторон. Упомянутая выше кость, заложившаяся в скелетогенной мезенхиме слезно-носового протока и ограничивающая его в месте разветвления медиально, образует в это время латерально выступающую пластинку, которая прикрывает среднюю часть протока с дорсальной стороны. В отличие от того, что имеется у аксолотля, дорсо-каудальный конец этой кости не заворачивает над крышей обонятельной капсулы, а лежит только латерально от нее, так как у T. taeniatus frontale сама дорастает до дорсо-латерального угла крыши. Таким образом передне-вентральный конец описываемой кости, так же как у аксолотля, занимает положение lacrymale низших хвостатых амфибий, а ее каудо-дорсальный конец - положение praefrontale. Хотя у обыкновенного тритона ни разу не отмечалась двойная закладка этой кости, принимаемой у высших хвостатых амфибий за praefrontale, все же ее развитие совершается тем же путем, что и у аксолотля, и сомнений в гомологии этой кости у аксолотля и обыкновенного тритона не возникает. Поскольку же варианты в ее развитии у первого указывают на двойственную природу кости и вызывают сомнения в правильности принятого для нее наименования, она в дальнейших описаниях будет обозначаться как praefrontale в кавычках.
В конце метаморфоза (личинки длиной 29 - 32 мм) в месте раздвоения протока и перед ним наружный конец латеральной пластинки передне-вентральной части "praefrontale" подходит к дорсальному концу восходящего отростка maxillare, а вентральный край "praefrontale", ограничивающий среднюю часть протока с медиальной стороны, подрастает к медиальному выступу этого же отростка maxillare, расположенному под протоком. Таким образом, обе эти кости окружают слезно-носовой проток практически полным кольцом в месте его перехода из средней части в заднюю. Идентичные отношения сохраняются и у взрослых животных (рис. 55), у которых средняя часть слезно-носового протока проходит в костном канале, образованном обеими костями: "praefrontale" и maxillare. Каудальная часть протока канализируется только в самом конце метаморфоза, включая обе веточки, которые открываются наружу друг над другом у внутреннего угла глаза. Очень редко, но может отсутствовать одна из ветвей; поскольку они оканчиваются в коже на одном уровне, определить, какая из них отсутствует, трудно. Один раз наблюдалась независимая закладка медиальной ветви. Septomaxillare у обыкновенного тритона отсутствует.
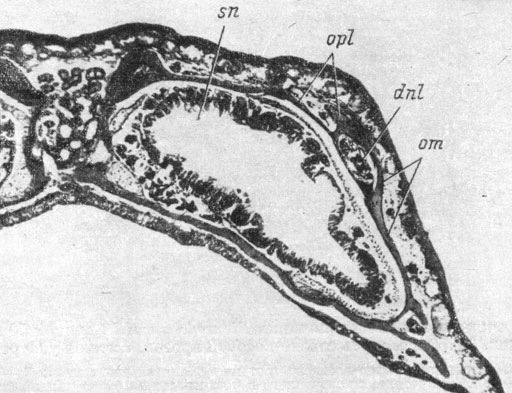
Рис. 55. Поперечный срез правой стороны головы взрослого Triton taeniatus длиной 56 мм, на уровне середины слезно-носового протока. (Увел. 40)
У T. cristatus karelini в отличие от предыдущего вида не наблюдается такого сокращенного развития закладки слезно-носового протока: дифференцировка ее идет более постепенно, хотя также сокращена по сравнению с ходом этого процесса у Hynobiidae. На стадии 58 (личинка длиной 23.5 мм) никакой видимой дифференцировки покровного эпителия еще нет, но под его участком, расположенным над местом отхождения якобсонова органа от обонятельного мешка, отмечается небольшое скопление мезенхимных клеток. На этой же стадии развития, но у более крупной личинки (длиной 26 мм) в этом месте покровный эпителий образует небольшую выпуклость в глубь тканей, вызванную концентрацией клеток базального слоя. Количество мезенхимы под местом закладки протока увеличилось. Такое же состояние сохраняется и на стадии 60 (личинка длиной 27 мм).
На стадии 61 (личинки длиной 29 - 31 мм) клетки базального слоя эпителия в переднем отрезке закладки протока группируются в плоский тяж, заметно вдающийся в глубь тканей. Кзади этот тяж расширяется в клеточный пласт. Вся закладка протока разрастается на длину протяжения якобсонова органа. К концу этой стадии развития передний конец закладки слезно-носового протока преобразуется в плотный округлый, уже спустившийся под покровный эпителий тяж, но лежащий еще над его базальной мембраной и далее кзади переходящий в клеточный пласт, на конце раздваивающийся. Никакого углубления наружной поверхности покровного эпителия в месте закладки протока у T. cristatus karelini не наблюдается.
На стадии 62 (личинка длиной 31 мм) передний конец тяжа слезно-носового протока начинает расти в глубь тканей, а на стадии 63 (личинка длиной 34 мм) он уже соединяется с латеральным дивертикулом обонятельного мешка и от полости дивертикула начинается образование полости в переднем отрезке протока. Слезно-носовой проток погружен в ткани на всем своем протяжении, за исключением каудальной, разветвленной части, веточки которой еще проходят внутрикожно, над базальной мембраной покровного эпителия.
В предметаморфозном периоде (личинка длиной 46 мм) у T. cristatus karelini закладывается кость, соответствующая lacrymale и praefrontale низших хвостатых амфибий. В отличие от развития "praefrontale" у аксолотля и обыкновенного тритона закладка этой кости начинается с ее каудо-дорсальной части, лежащей вдоль латеральной стороны хряща обонятельной капсулы, сзади от разветвления слезно-носового протока (т. е. с части, соответствующей истинной praefrontale низших хвостатых амфибий), и разрастающейся сначала каудально и дорсально. В это время maxillare уже разрослась дорсально, образовав восходящий отросток, и его верхний край прикрывает с латеральной стороны среднюю часть и передний отрезок каудальной части слезно-носового протока. Образование полости последнего может начинаться и не от полости латерального дивертикула обонятельного мешка, но все же в передней (проксимальной) части протока.
В начале метаморфоза (личинка длиной 55 мм) "praefrontale" разрастается вперед, ограничивая каудальную часть слезно-носового протока с медиальной стороны, и образует латерально выступающую пластинку, которая прикрывает сверху конец средней части протока и место его разветвления, т. е. представляет собой ту часть кости, которая соответствует lacrymale низших хвостатых амфибий и с которой начинается ее развитие у аксолотля и обыкновенного тритона. Латеральный конец этой пластинки близко подходит к дорсальному краю восходящего отростка maxillare. В своей каудальной части "praefrontale" разрастается еще более дорсально и над латеро-дорсальным углом крыши обонятельной капсулы смыкается с frontale. Слезно-носовой проток канализируется полностью в середине метаморфоза. К концу метаморфоза (личинка длиной 61 мм) "praefrontale" и восходящий отросток maxillare окружают проток в конце его средней части и в месте разветвления так же, как у обыкновенного тритона, практически полным кольцом, причем первая образует дорсальную и медиальную поверхности этого кольца, а второй - латеральную и вентральную. Восходящий отросток maxillare продолжается вперед и прикрывает с латеральной стороны также всю среднюю и каудальный конец передней части слезно-носового протока.
После окончания метаморфоза проток открывается у угла глаза, на внутренней поверхности нижнего века, как правило, двумя ветвями. Но иногда одна из ветвей может отсутствовать, а могут образовываться и три ветви. Наблюдается также и независимая закладка отдельных веточек и вторичное соединение их с протоком. Septomaxillare у T. cristatus karelini отсутствует, у обычного гребенчатого тритона - T. cristatus - ее тоже нет.
Развитие и редукция слезно-носового протока у Pleurodeles waltlii. Хотя испанский тритон - P. waltlii - относится к сем. Salamandridae, описание развития у него слезно-носового протока выделено в самостоятельный раздел, поскольку оно сопровождается у этого вида, практически полностью ведущего водный образ жизни, весьма своеобразным процессом последующей редукции протока.
Развитие слезно-носового протока у P. waltlii (стадии развития везде указываются по таблицам Гальена и Дюроше (Gallien et Durocher, 1957)) начинается на стадиях 52 - 53 (личинки 24 - 27.5 мм длиной) с утолщения в покровном эпителии, расположенного между ноздрей и глазом, над местом отхождения якобсонова органа от обонятельного мешка. В дальнейшем это, вызвайное концентрацией клеток базального слоя, внутри-кожное утолщение, под которым наблюдается небольшое скопление мезенхимы, превращается в эпителиальный тяж, передний конец которого значительно не доходит до ноздри, заканчиваясь примерно на уровне середины обонятельного мешка (стадии 53 - 54, личинки длиной 30 - 31.5 мм). Каудальный конец тяжа слезно-носового протока не достигает глаза и находится на уровне начала хоаны, он значительно шире переднего конца и в нем заметно намечающееся раздвоение. Затем этот тяж в своей средней части отделяется от покровного эпителия и погружается под него, но идет еще близко от поверхности, оставаясь подостланным пограничной базальной мембраной. Передний и каудальный концы тяжа протока остаются соединенными с покровным эпителием, причем каудальный конец закладки разделяется на две ветви. Передний конец закладки протока по-прежнему не достигает ноздри (стадии 54 - 55а - начало метаморфоза, личинки длиной 36 - 38 мм).
Несколько позже (стадии 55а - 55b - середина метаморфоза, личинки длиной 40 - 48 мм) передний конец тяжа, который теперь лучше называть проксимальным (а каудальный соответственно дистальным), отделяется от покровного эпителия, начинает расти в глубь тканей, подрастает к латеральному дивертикулу обонятельного мешка и срастается с ним. В тяже за счет расхождения составляющих его клеток образуется полость, сначала в средней, затем в его проксимальной и дистальной частях, и он превращается в канал, открывающийся наружу двумя ветвями у края нижнего века. В качестве вариации может наблюдаться отсутствие одной из ветвей, а также ее независимая закладка.
На стадии 55а в мезенхиме, окружающей каудальный конец средней части слезно-носового протока и место его разветвления, медиально от протока - между ним и хрящом обонятельной капсулы - образуется кость, доходящая кзади до уровня впадения ветвей протока в кожу. Эта кость, принимаемая у высших хвостатых амфибий за praefrontale, закладывается у P. waltlii на месте, типичном для lacrymale низших хвостатых амфибий, так же как у аксолотля. Дорсальный конец восходящего отростка maxillare в это время еще намного не доходит до вентральной стороны протока. На стадии 55b восходящий отросток maxillare уже настолько разрастается дорсально, что ограничивает слезно-носовой проток с латеральной стороны почти на всем его протяжении, начиная с его переднего (проксимального) конца в месте впадения в латеральный дивертикул обонятельного мешка. В каудальном отделе средней части протока восходящий отросток maxillare подстилает проток и с вентральной стороны за счет образовавшегося медиального выступа. С медиальной стороны средняя часть слезно-носового протока ограничивается передней частью "praefrontale", вентральный край которой начинает срастаться под протоком с медиальным выступом восходящего отростка maxillare. Дорсальная сторона средней части протока прикрывается латеральной пластинкой передней части "praefrontale". Таким образом, средняя часть слезно-носового протока проходит в канале, составленном обеими костями.1
1 (У некоторых закончивших метаморфоз особей передний отдел средней части слезно-носового протока в случае его сохранения может проходить в костном канале, находящемся целиком внутри восходящего отростка maxillare.)
В своей каудальной части "praefrontale" разрастается дорсально вдоль хряща обонятельной капсулы, занимая место типичной praefrontale низших хвостатых амфибий, и над латеро-дорсальным углом крыши капсулы смыкается с frontale, налегая на ее латеральный край. Septomaxillare у P. waltlii не образуется. Таким образом, развитие слезно-носового протока и связанных с ним костей не имеет у P. waltlii принципиальных отличий от этого процесса у других представителей сем. Salamandridae и у аксолотля. Стоит отметить, что сроки закладки и развития слезно-носового протока у P. waltlii могут значительно варьировать, но, как правило, к середине метаморфоза проток оказывается полностью развитым.
Однако у испанского тритона имеет место также и обратное развитие слезно-носового протока, которое начинается вскоре после его образования (стадии 55b - 55с, середина - конец метаморфоза). Нужно сказать, что редукция слезно-носового протока у этого вида - процесс, явно не установившийся, и наблюдается далеко не у каждой особи. Не является фиксированным также и момент начала редукции: в тех случаях, когда этот процесс начинается поздно (иногда после окончания метаморфоза), свидетельством предыдущего существования слезно-носового протока остается окружавший его костный канал, каудо-медио-дорсальная половина которого образуется за счет "praefrontale", а ростро-латеро-вентральная - за счет восходящего отростка maxillare. В случае раннего начала редукции слезно-носового протока канал не успевает образоваться, и тогда дорсальный конец восходящего отростка maxillare просто налегает на вентральный край "praefrontale". Встречаются также взрослые особи, у которых на одной стороне имеется слезно-носовой проток, а на другой он редуцирован (рис. 56, А, Б). Возможно и длительное сохранение дистальной части протока при полном отсутствии проксимальной. Все эти факты показывают, что имеющийся еще в настоящее время у P. waltlii слезно-носовой проток, вероятно, в связи с преимущественно водным образом жизни этого вида, в недалеком будущем будет утрачен.
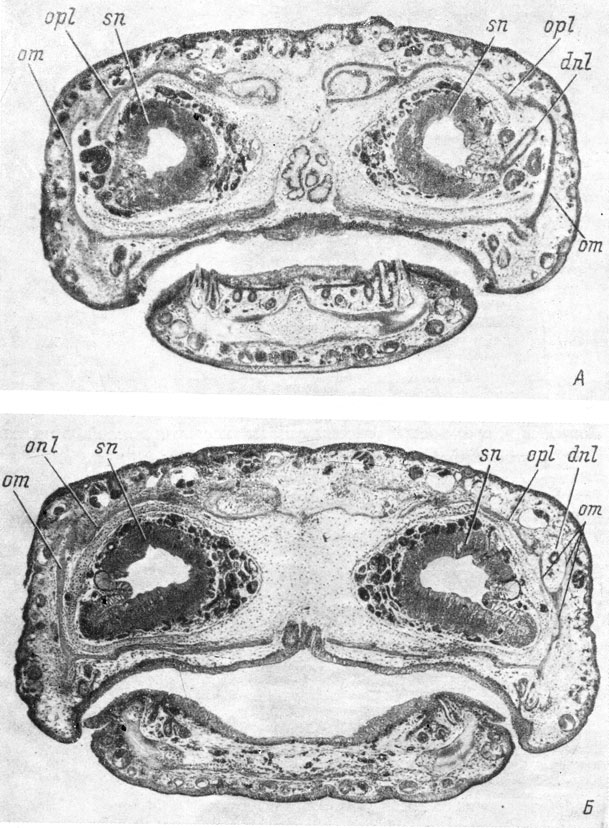
Рис. 56. Поперечные срезы головы личинки Pleurodeles waltlii длиной 50 мм, конец метаморфоза. (Увел. 36). Справа есть слезно-носовой проток, слева он уже редуцировался. А - срез на уровне впадения слезно-носового протока в обонятельный мешок; Б - срез на уровне середины слезно-носового протока, заключенного в костный канал (слева канала нет)
Редукция слезно-носового протока всегда начинается с его проксимального конца, и особый интерес представляют начальные стадии этого процесса, так как при этом проксимальный конец протока начинает передвигаться по боковой стенке обонятельного мешка по направлению к наружному носовому отверстию. В это время (стадии 55b - 55с, середина-конец метаморфоза) можно видеть проксимальный (передний) конец протока присоединенным к стенке вводного канала органа обоняния (рис. 57, Б), к краю ноздри (рис. 57, А) и, наконец, оканчивающимся в коже около заднего края ноздри. В то же время на другой стороне зачастую имеется слезно-носовой проток, впадающий, как ему и полагается, в латеральный дивертикул обонятельного мешка (рис. 57, В). Следовательно, в начале процесса редукции проксимальный конец слезно-носового протока P. waltlii проделывает в обратном направлении тот же самый путь, который прослеживается при нормальном развитии протока у низших хвостатых амфибий сем. Hynobiidae, a Ambystomidae и Salamandridae, в том числе и у самого испанского тритона, оказался утраченным вследствие укорочения процесса развития и выпадения отдельных его стадий.
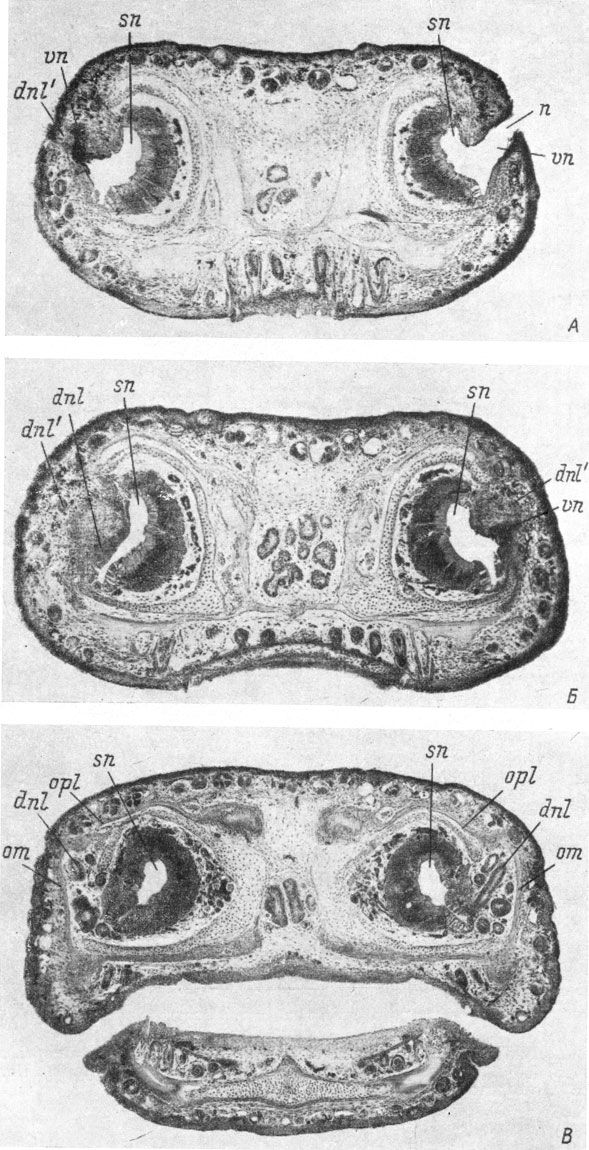
Рис. 57. Поперечные срезы головы личинки Pleurodeles waltlii длиной 48 мм, конец метаморфоза. (Увел. 36). На обеих сторонах имеется дополнительный передний конец слезно-носового протока. Справа первоначальный передний конец протока сохранился; слева он уже редуцирован. А - срез на уровне ноздрей (слева - дополнительный передний конец протока открывается у заднего края ноздри); Б - срез, проходящий несколько сзади ноздрей (слева - дополнительный передний конец протока и редуцирующийся остаток его первоначального переднего конца, продвинувшегося по боковой стенке обонятельного мешка вперед; справа - дополнительный передний конец протока, впадающий в стенку вводного канала); В - срез на уровне впадения слезно-носового протока в обонятельный мешок (справа - еще не редуцированный первоначальный передний конец протока)
Лишь после описанного последовательного передвижения проксимального (переднего) конца слезно-носового протока по боковой стенке обонятельного мешка и стенке вводного канала к краю ноздри и затем к покровному эпителию начинается резорбция этого переднего конца, в ряде случаев распространяющаяся и на дистальную (каудальную) половину протока и приводящая в результате к его полному исчезновению (рис. 56). При более позднем начале редукции (в самом конце метаморфоза и после его окончания) передвижения проксимального (переднего) конца слезно-носового протока не происходит, так как этому процессу мешает фиксированное положение протока внутри костного канала. В этих случаях образуется дополнительный проксимальный (передний) конец слезно-носового протока, который также последовательно присоединяется к стенке вводного канала, к краю ноздри, а затем к покровному эпителию (рис. 57, А, Б). После образования и постепенного изменения места присоединения дополнительного переднего конца протока первоначальный проксимальный его конец, впадавший в латеральный дивертикул обонятельного мешка, редуцируется (рис. 57, Б), но какое-то время оба эти конца сосуществуют. Редуцируется и вновь образовавшийся дополнительный передний конец. Следовательно, и в случаях более позднего наступления процесса редукции слезно-носового протока, когда обратному перемещению его первоначального проксимального конца препятствует уже образовавшийся костный канал, все же воспроизводится в обратном порядке исходный путь передвижения переднего конца протока, только этот путь повторяется не первоначальным, а его дополнительно образующимся проксимальным концом.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'