Обсуждение результатов экспериментов
Эксперименты по частичному удалению обонятельной плакоды, оказавшиеся результативными, были проведены в общей сложности у 447 эмбрионов амфибий, причем исследовались представители различных семейств, относящихся к низшим хвостатым амфибиям - Hynobiidae, высшим хвостатым амфибиям - Salamandridae, низшим бесхвостым амфибиям - Discoglossidae и высшим бесхвостым амфибиям - Ranidae, Полученные результаты дают материал для сравнения в эволюционном аспекте.
В табл. 8 показаны количественные соотношения случаев с образованием дефектного органа обоняния после удаления различных половин обонятельной плакоды и то же самое отдельно для якобсонова органа. При анализе данных этой таблицы бросаются в глаза две отчетливо выраженные закономерности.

Таблица 8. Результаты частичного удаления обонятельной плакоды у хвостатых и бесхвостых амфибий
Первая из этих закономерностей заключается в том, что при удалении по отдельности различных четырех половин обонятельной плакоды основное количество случаев с образованием дефектного органа обоняния приходится лишь на две удаляемые половины, причем у низших хвостатых, низших бесхвостых и высших бесхвостых амфибий - это нижняя и задняя половины плакоды, а у высших хвостатых амфибий - это задняя и верхняя половины плакоды. Вторая наблюдаемая закономерность состоит в том, что наибольшее количество случаев с отсутствием или дефектностью якобсонова органа в каждой из исследованных четырех групп амфибий приходится на серии с удалением тех же самых половин обонятельной плакоды. Соответственно у низших хвостатых, низших бесхвостых и высших бесхвостых амфибий местом локализации презумптивного материала ноздри, вводного канала, хоанального канала и якобсонова органа является квадрант III, общий для нижней и задней половин обонятельной плакоды, а у высших хвостатых амфибий местом локализации этого материала служит квадрант IV, общий для задней и верхней ее половин (рис. 44).
Отдельные случаи возникновения дефектного органа обоняния при удалении тех половин обонятельной плакоды, в которых, судя по результатам экспериментов, нет соответствующего материала, объясняются либо случайным захватом участка квадранта, содержащего презумптивный материал указанных отделов органа обоняния, либо некоторым распространением этого материала за границы условно обозначенного квадранта. Эти случаи позволяют уточнить локализацию материала ноздри и вводного канала, с одной стороны, и хоанального канала - с другой. Например, у Salamandridae, помимо случаев дефектности органа обоняния, возникших после удаления квадранта IV, имеются еще два случая образования дефектного органа обоняния: один при удалении передней и один при удалении нижней половины обонятельной плакоды. Случаи эти различны по результатам: при удалении нижней половины плакоды нет ноздри и поврежден вводной канал, при удалении передней половины ноздря и вводной канал нормальны, но зато отсутствует хоанальный канал. Следовательно, материал ноздри находится в квадранте IV на границе с нижней половиной плакоды, т. е. на границе с квадрантом III, а материал хоанального канала - на границе с передней половиной, т. е. на границе с квадрантом I (рис. 44).
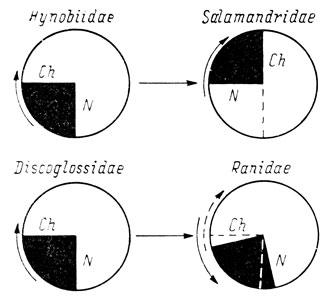
Рис. 44. Схема расположения презумптивного материала ноздри и хоаны в обонятельной плакоде амфибий. Стрелками показано направление поворота плакоды
У низших хвостатых амфибий (Hynobiidae) дефектны ноздря и вводной канал в 2 случаях удаления передней половины обонятельной плакоды, а хоанальный канал отсутствует в 1 случае удаления верхней половины. Соответственно презумптивный материал ноздри располагается в квадранте III на границе с передней половиной плакоды, т. е. на границе с квадрантом II, а материал хоанального канала - на границе с верхней половиной, т. е. на границе с квадрантом IV (рис. 44).
У низших бесхвостых амфибий (Discoglossidae) наблюдаются совершенно те же соотношения, что у Hynobiidae: случаи, имеющиеся при удалении передней половины обонятельной плакоды, касаются ноздри и вводного канала, случай при удалении верхней половины плакоды связан с отсутствием хоанального канала. Следовательно, у низших хвостатых и бесхвостых амфибий материал ноздри и вводного канала расположен одинаково в квадранте III на границе с квадрантом II. Так же одинаково локализован у них и материал хоанального канала: в том же квадранте III, но на границе с квадрантом IV (рис. 44).
У высших бесхвостых амфибий Ranidae презумптивный материал ноздри, вводного канала, хоанального канала и якобсонова органа располагается так же, как у низших амфибий в квадранте III с одним небольшим изменением: весь этот комплекс материала, по-видимому, немного сдвинут в сторону квадранта II, о чем свидетельствуют наличие 4 случаев дефектности вводного канала и 5 случаев дефектности якобсонова органа после удаления передней половины обонятельной плакоды и отсутствие случаев повреждения хоанального канала после удаления ее верхней половины, а также безусловное преобладание случаев с дефектностью ноздри и вводного канала (в общей сложности 1-5) при удалении нижней половины плакоды, в которую входят оба квадранта III и II, по сравнению с удалением задней половины, в которую квадрант II не входит и где был всего один случай дефектности ноздри и вводного канала.
На схеме (рис. 44) изображена локализация презумптивного материала ноздри и хоаны у всех четырех групп амфибий. У низших амфибий, как у хвостатых, так и у бесхвостых, локализация презумптивного материала ноздри, вводного канала, хоанального канала и якобсонова органа совершенно одинакова как по положению его в обонятельной плакоде, так и по соотношению материала ноздри и вводного канала, с одной стороны, и хоанального канала - с другой. То же самое касается и высших бесхвостых амфибий с поправкой на небольшой сдвиг всего комплекса материала на территорию квадранта II. Если же сравнить локализацию этого материала с локализацией его у высших хвостатых амфибий, то видно, что расположение материала ноздри и хоанального канала по соотношению друг с другом остается тем же самым, но весь этот комплекс оказывается сдвинутым из квадранта III в квадрант IV. Другими словами, соотношение презумптивного материала ноздри и хоанального канала в обонятельной плакоде одинаково у всех амфибий, но сама плакода у высших хвостатых амфибий повернута, примерно, на 90° по часовой стрелке по сравнению с низшими амфибиями, а также с высшими бесхвостыми. Соответственно передвинут и материал якобсонова органа, входящий в этот комплекс.
Следовательно, филогенетический поворот органа обоняния предков амфибий зафиксирован не только в сдвиге во время эмбрионального развития закладки якобсонова органа с медиальной стороны на латеральную и повороте обонятельного мешка, но и в повороте самой обонятельной плакоды. В развитии высших хвостатых амфибий стадия первоначального положения материала указанного комплекса, включая якобсонов орган, в квадранте III обонятельной плакоды уже выпала, и только исследование более примитивных амфибий помогает установить наличие такого поворота. В развитии высших бесхвостых амфибий, по-видимому, выпала не только первоначальная стадия локализации материала в квадранте III, но и последующая стадия локализации этого материала в квадранте IV, а его настоящее положение в квадранте III следует считать вторичным, а не первичным. Выше говорилось (стр. 58 - 59), что у высших бесхвостых амфибий отмечается первоначальный сдвиг закладки якобсонова органа, расположенной исходно медиально, в латеральную сторону, так же как это имеет место у хвостатых амфибий и у Bombina, но этот сдвиг быстро прекращается, и начинается обратное перемещение якобсонова органа в медиальном направлении, в результате чего конечное положение органа оказывается более медиальным, чем первоначальное. Отсюда был сделан вывод, что медиальное положение якобсонова органа у высших бесхвостых амфибий вторично. Соответственно этому и произошло, по-видимому, в ряду бесхвостых амфибий обратное перемещение материала из промежуточного положения в квадранте IV к исходному в квадранте III, т. е. точнее произошел обратный поворот обонятельной плакоды. Однако плакода не зафиксировалась в этом исходном положении, а повернулась немного дальше, благодаря чему рассматриваемый материал частично сдвинулся из квадранта III в квадрант II, и поэтому его положение и не является вполне идентичным положению у низших амфибий. Таким образом, данные по нормальному развитию якобсонова органа и экспериментальные данные о локализации презумптивного материала в обонятельной плакоде хорошо согласуются и вместе с тем дополняют друг друга, подтверждая вывод о вторичности медиального положения якобсонова органа, наблюдающегося у большинства бесхвостых амфибий. В то же время одинаковая локализация презумптивного материала якобсонова органа у низших хвостатых и низших бесхвостых амфибий служит дополнительным доказательством гомологии этого образования у обоих отрядов.
Необходимо еще обратить внимание на локализацию презумптивного материала ноздри и вводного канала, с одной стороны, и хоанального канала - с другой, по отношению друг к другу. Фактически материал этих отделов органа обоняния располагается вместе, в непосредственной близости друг от друга, в одном квадранте обонятельной плакоды, хотя в дефинитивном органе ноздря и хоана находятся на диаметрально противоположных его концах. Отмеченный факт имеет огромное значение для установления происхождения хоан наземных позвоночных и будет подробно рассмотрен в дальнейшем при обсуждении этого вопроса. Другим фактом, имеющим не меньшее значение для затронутого вопроса, является локализация материала самого хоанального канала в обонятельной плакоде низших амфибий. Презумптивный материал хоанального канала располагается у примитивных амфибий в верхней стороне квадранта III, на границе с нижней стороной квадранта IV (рис. 44), и именно в этом месте к обонятельной плакоде подходит закладка инфраорбиталь-ной сейсмосенсорной линии (см. рис. 41, А). Значение этого обстоятельства будет также разобрано ниже при обсуждении вопроса о происхождении хоан (глава VII).
Следует еще остановиться на имевших место после частичного удаления обонятельной плакоды нарушениях развития самой хоаны, образующейся, как было показано выше при описании нормального развития и результатов экспериментов с полной экстирпацией плакоды, не из ее материала, а из материала ротовой полости. Эти нарушения развития хоаны заключались в недоразвитии хоанальных клапанов и в их сращении друг с другом. Что касается последнего, то такой дефект следует отнести к дефектам развития хоанального канала, поскольку при сращении клапанов нарушается связь между органом обоняния и хоаной, которую осуществляет хоанальный канал, и тем самым сквозное соединение с ротовой полостью.
Случаи с отсутствием и уменьшением хоанального клапана у Hynobiidae наблюдались только у личинок, зафиксированных в начале метаморфоза, во время которого в норме хоанальный клапан резорбируется; вследствие операционного вмешательства этот процесс на оперированной стороне мог несколько ускориться. Поэтому эти случаи не противоречат результатам, полученным после полного удаления обонятельной плакоды у Hynobiidae, когда образовывалась нормальная хоана с нормальным хоанальным клапаном (стр. 73), и не снимают сделанного вывода, что даже отсутствие органа обоняния в целом не влияет на образование нормальной хоаны у примитивных хвостатых амфибий.
В отношении высших хвостатых, низших бесхвостых и высших бесхвостых амфибий дело обстоит по-другому. Все случаи отсутствия и дефектности хоанальных клапанов у этих групп связаны с отсутствием или дефектностью хоанального канала. Это обстоятельство доказывает необходимость контакта хоанального канала с хоаной для осуществления нормального развития клапанов. Данные по нормальному развитию хоаны у бесхвостых амфибий показывают, что в построении медиального хоанального клапана участвует материал хоанального канала (стр. 48), и эти данные полностью подтверждаются отсутствием или дефектностью медиального клапана у Bombina во всех случаях отсутствия хоанального канала. У Rana к этому присоединяется и неправильное развитие латерального клапана. Возможно, что у высших бесхвостых амфибий материал хоанального канала начал принимать участие в построении и латерального клапана хоаны, но в равной мере возможно и нарушение индукционного формативного взаимодействия вследствие отсутствия непосредственного контакта между хоанальным каналом и хоаной. Ослабление индуктивного взаимодействия безусловно играет роль в нарушении развития медиального хоанального клапана после полного удаления обонятельной плакоды (см. стр. 69).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'