Бесхвостые амфибии
Discoglossidae. Из представителей этого семейства экспериментам подверглась Bombina bombina. Операции производились на эмбрионах размером от 4.5 мм (стадия поздней хвостовой почки) до 6 мм (стадия образования жаберных выростов). Поскольку у бесхвостых амфибий уже на самых ранних стадиях развития иная форма головы и иное расположение обонятельной плакоды, чем у хвостатых, то для возможности последующего сравнения результатов в основу бралось ее положение по отношению к глазу. При этом обращенная к глазу половина обонятельной плакоды бесхвостых, так же как и у хвостатых амфибий, условно получила наименование верхней, хотя по своему расположению у первых она верхней не является (рис. 43, А, Б). Остальные половины обонятельной плакоды обозначены теми же названиями, что и у хвостатых амфибий, соответственно их отношению к половине, принимаемой за верхнюю.
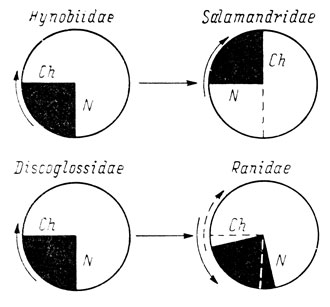
Рис. 43. Схема операций удаления по частям обонятельной плакоды бесхвостых амфибий. А - положение обонятельной плакоды в момент операции; Б - увеличенная схема обонятельной плакоды с обозначением удалявшихся половин и квадрантов
Подопытные личинки фиксировались на разных стадиях развития, начиная с момента образования хоан и якобсонова органа и кончая предметаморфозными. Всего было прооперировано 105 эмбрионов, выращено, зафиксировано и исследовано микроскопически 93 личинки. Препараты окрашивались по Маллори. По сериям опытов оперированные личинки распределяются следующим образом: 1-я серия (удаление передней половины обонятельной плакоды) - 26, 2-я серия (удаление нижней половины плакоды) - 23, 3-я серия (удаление задней половины плакоды) - 25 и 4-я серия (удаление верхней половины плакоды) - 19 экз. Результаты исследования оказались следующими.
В 1-й серии (удаление передней половины обонятельной плакоды) из 26 подопытных личинок орган обоняния оказался нормально развитым у 21, в 9 из этих случаев он был уменьшен. Дегенерирующих органов обоняния не было. Дефектный орган обоняния наблюдался у 5 экз., причем среди них не было ни одного случая отсутствия или дефектности хоанального канала и якобсонова органа. Дефектность обонятельного органа осуществлялась только за счет дефектности ноздри (2 случая) и дефектности вводного канала (4 случая). Кроме того, в 2 случаях отмечены недоразвитие и деформация главной обонятельной полости.
Во 2-й серии опытов (удаление нижней половины обонятельной плакоды) из 23 подопытных личинок 12 имели нормальный орган обоняния, в 4 из этих случаев уменьшенный. 11 личинок обладали дефектным органом обоняния. Случаев с дегенерацией органа не было. Дефектность органа обоняния выразилась в отсутствии якобсонова органа (7 случаев) или в его дефектности (3 случая), в отсутствии хоанального канала (2 случая), дефектности ноздри (5 случаев) и вводного канала (5 случаев). Кроме того, имелись случаи нарушения развития самой хоаны, заключавшиеся в отсутствии медиального хоанального клапана (2 случая) или в его недоразвитии (1 случай), а также в сращении клапанов друг с другом (1 случай). Все случаи отсутствия или дефектности медиального клапана хоаны связаны с отсутствием или дефектностью хоанального канала. При сращении клапанов друг с другом хоанальный канал на всем своем остальном протяжении был развит нормально, но само сращение клапанов означает отсутствие связи полости хоанального канала с ротовой полостью и может быть приравнено к дефектности каудального отрезка этого канала. В 2 случаях выявилось неправильное распределение обонятельного эпителия в главной полости и уменьшение его количества.
В 3-й серии экспериментов (удаление задней половины обонятельной плакоды) 9 из 25 оперированных личинок обладали нормально развитым органом обоняния, уменьшенных органов обоняния не было. В 1 случае обонятельный орган находился в состоянии дегенерации, и в 15 случаях он был дефектным. В последних 15 случаях дефектность органа обоняния обусловливалась отсутствием якобсонова органа (9 случаев) или его дефектностью (3 случая), отсутствием хоанального канала (12 случаев) или его дефектностью (1 случай), дефектностью ноздри (4 случая) и вводного канала (3 случая). Наблюдались также отсутствие или дефектность медиального клапана хоаны - все 15 случаев, из них в 3 случаях клапаны срослись друг с другом, т. е. было нарушено развитие каудального конца хоанального канала. В единственном случае наличия дегенерирующего органа обоняния в виде комка клеток, не соединенного с хоаной, медиальный клапан тоже был недоразвит, так что общее число случаев с нарушением развития медиального хоанального клапана составляет 16. Кроме того, имелись 9 случаев с неправильным распределением и уменьшенным количеством обонятельного эпителия в главной полости, а также в 3 случаях наблюдались недоразвитие и деформация этой полости.
В 4-й серии опытов (удаление верхней половины обонятельной плакоды) 17 из 19 оперированных личинок имели нормальный орган обоняния, в 4 из этих случаев он был немного уменьшен. В 1 случае орган обоняния отсутствовал полностью, за исключением хоаны, имевшей только латеральный клапан. Дегенерирующих органов обоняния не было. У 1 личинки орган обоняния оказался дефектным за счет отсутствия хоанального канала, неправильного распределения обонятельного эпителия главной полости и отсутствия медиального клапана хоаны.
Из сказанного видно, что основные результаты частичного удаления обонятельной плакоды у B. bombina получились такие же, как у высших хвостатых амфибий: нормальное развитие органа обоняния, развитие дефектного органа обоняния, его полное отсутствие и дегенерация. Здесь необходимо отметить, что при определении нормально развитых органов обоняния и при подсчете этих случаев не учитывалось отсутствие наиболее рано развивающегося добавочного мешка - латерального аппендикса, поскольку это образование существует у B. bombina очень недолго (со стадии 8.5 мм до стадии 12 - 13 мм) и быстро подвергается резорбции. Так как фиксация подопытного материала в основном производилась на гораздо более поздних стадиях развития, то учесть в данных случаях отсутствие латерального аппендикса не было возможности. Прибавление же случаев с отсутствием латерального аппендикса при фиксации личинок на тех стадиях развития, на которых он еще имеется, к числу случаев с дефектным органом обоняния могло только спутать общую картину соотношения количества нормальных и дефектных органов обоняния, не давая вместе с тем достоверных данных для сравнения.
Количественное соотношение результатов операций показано в табл. 6.
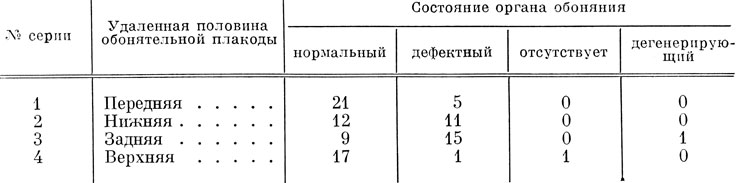
Таблица 6. Результаты частичного удаления обонятельной плакоды у Discoglossidae
При анализе данных, приведенных в табл. 6, прежде всего бросается в глаза, что несмотря на наличие 1 случая отсутствия органа обоняния, чего не было у Hynobiidae, общее число случаев отсутствия или дегенерации органа обоняния у B. bombina, как и у низших хвостатых амфибий, значительно меньше, чем у высших Urodela, - их всего 2 на 93 подопытных экземпляра, и они вследствие своей малочисленности практического значения не имеют. В отношении же личинок с нормально развитым и дефектным органом обоняния наблюдается, как и у хвостатых амфибий, явно выраженная закономерность: 26 из имеющихся 32 случаев наличия дефектного органа обоняния приходится на две экспериментальные серии из четырех - серию с удалением нижней и серию с удалением задней половин обонятельной плакоды, т. е. у Discoglossidae выявляются совершенно те же соотношения, которые наблюдались у низших хвостатых амфибий (Hynobiidae), и, следовательно, презумптдвный материал ноздри, вводного канала, хоанального канала и якобсонова органа локализуется так же, как у последних, в квадранте III, являющемся общим для нижней и задней половин обонятельной плакоды.
6 остальных случаев с наличием дефектного органа обоняния, из которых 5 приходится на серию с удалением передней половины плакоды и 1 случай - на серию с удалением верхней половины плакоды, опять-таки, как и в опытах над хвостатыми амфибиями, дают возможность более точного установления локализации презумптивного материала ноздри и вводного канала, с одной стороны, и хоанального канала - с другой. Во всех 5 случаях, падающих на серию с удалением передней половины обонятельной плакоды, дефектность органа обоняния выражалась в дефектности ноздри и вводного канала, а хоанальный канал во всех этих случаях был развит нормально. Напротив, в 1 случае дефектности органа обоняния, относящемся к серии с удалением верхней половины плакоды, отсутствовал хоанальный канал, но зато ноздря и вводной канал были в полном порядке. Отсюда ясно, что презумптивный материал ноздри и вводного канала располагается в квадранте III на границе с передней половиной обонятельной плакоды, т. е. на границе с квадрантом П. В то же время материал хоанального канала лежит в том же квадранте III на границе с верхней половиной плакоды, т. е. на границе его с квадрантом IV. Оказывается, что точная локализация презумптивного материала ноздри, вводного канала и хоанального канала у B. bombina совершенно такая же, как это имеет место у низших хвостатых амфибий.
У B. bombina при частичном удалении обонятельной плакоды, помимо возникновения дефектности органа обоняния за счет ноздри, вводного канала, хоанального канала и якобсонова органа, выявились также нарушения развития, относящиеся к неправильному распределению обонятельного эпителия и уменьшению его количества в главной носовой полости, а также к недоразвитию и наличию деформации этой полости при сохранении ее связи с внешней средой. Поскольку эти нарушения развития главной носовой полости имели место во всех 4 сериях экспериментов, можно думать, что презумптивный материал этой полости либо рассредоточен по всей обонятельной плакоде, либо локализован в середине ее, и поэтому удаление любой половины плакоды способно отразиться на развитии главной полости органа обоняния.
Кроме того, у B. bombina так же, как и у хвостатых амфибий, наблюдались нарушения в развитии хоаны, хотя она образуется не из материала обонятельной плакоды. Эти нарушения выразились в отсутствии или недоразвитии медиального хоанального клапана, а также в сращении обоих клапанов друг с другом. Все случаи нарушения развития медиального клапана хоаны имели место только при одновременном отсутствии хоанального канала и хорошо согласуются как с данными по нормальному развитию хоанального канала у бесхвостых амфибий (стр. 48), так и с результатами полного удаления обонятельной плакоды (стр. 69), показывая, что для нормального развития медиального клапана хоаны необходимо также и участие материала хоанального канала, т. е. необходимо соединение последнего с ротовой полостью.
Ranidae. В качестве подопытного материала из представителей этого семейства бесхвостых амфибий была использована Rana esculenta. Операции производились на эмбрионах длиной от 4 мм (стадия средней хвостовой почки) до 6 мм (стадия образования жаберных выростов). Всего было прооперировано 154 эмбриона, из них выращено, зафиксировано и исследовано 117 личинок. Фиксация проводилась на различных стадиях развития, начиная с момента образования хоан и якобсонова органа и до начала метаморфоза. Препараты окрашивались по Маллори. По отдельным сериям операций личинки распределились следующим образом: 1-я серия (удаление передней половины обонятельной плакоды) - 27, 2-я серия (удаление нижней половины плакоды) - 31, 3-я серия (удаление задней половины плакоды) - 30 и 4-я серия (удаление верхней половины плакоды) - 29 личинок. Поскольку по приведенным выше основаниям (стр. 84) у представителя низших бесхвостых амфибий - Bombina bombina - при определении числа личинок с нормальным и дефектным органом обоняния отсутствие латерального аппендикса не учитывалось, пришлось поступить таким же образом и при подсчете результатов, полученных в опытах над Rana esculenta, хотя у Rana он существует значительно дольше, резорбируется только во время метаморфоза, и соответственно у достаточно большого числа подопытных личинок орган обоняния не имел латерального аппендикса в результате операций, а не в результате его ранней резорбции, как у Bombina bombina. Однако при ином способе действий результаты, полученные у низших и высших бесхвостых амфибий, оказались бы несопоставимыми. Все же данные об отсутствии латерального аппендикса у Rana esculenta будут приведены отдельно и использованы для определения локализации его презумптивного материала. У R. esculenta результаты частичного удаления обонятельной плакоды оказались следующими.
В 1-й серии опытов (удаление передней половины обонятельной плакоды) у 20 из 27 оперированных личинок развился нормальный орган обоняния (без учета отсутствия латерального аппендикса). В остальных 7 случаях орган обоняния оказался дефектным за счет дефектности вводного канала (4 случая), дефектности якобсонова органа (5 случаев) и неправильного распределения обонятельного эпителия главной носовой полости (3 случая). Случаев дегенерации органа обоняния не было. Латеральный аппендикс отсутствовал в 15 случаях.
Во 2-й серии экспериментов (удаление нижней половины обонятельной плакоды) из 31 подопытной личинки 8 имели нормально развитый орган обоняния, а 23 - дефектный. Дефектность органа обоняния проявилась за счет дефектности ноздри (4 случая) и вводного канала (11 случаев), за счет якобсонова органа (4 случая полного отсутствия и 16 случаев дефектности), неправильного распределения обонятельного эпителия главной полости (3 случая), а также дефектности и деформации самой главной полости (3 случая). Как и в 1-й серии опытов, здесь не было ни одного случая нарушения развития хоанального канала, также не было случаев дегенерации органа обоняния. Не оказалось и случаев отсутствия латерального аппендикса.
В 3-й серии (удаление задней половины обонятельной плакоды) из 30 оперированных личинок у 19 образовался нормальный орган обоняния (без учета отсутствия латерального аппендикса), а 11 обладали дефектным органом. Дефектность органа обоняния осуществилась за счет дефектности ноздри и вводного канала (1 случай), за счет хоанального канала (2 случая отсутствия и 7 случаев дефектности), якобсонова органа (1 случай отсутствия и 3 случая дефектности), а также за счет дефектности и деформации главной полости. Кроме того, в 2 случаях наблюдалось неправильное развитие обоих хоанальных клапанов, как медиального, так и латерального, а в 2 случаях хоанальные клапаны частично срослись, сужая хоану до небольшого отверстия. Ни одного случая дегенерации органа обоняния не было. У 5 экз. отсутствовал латеральный аппендикс.
В 4-й серии опытов (удаление верхней половины обонятельной плакоды) все 29 оперированных личинок имели нормально развитый орган обоняния (без учета отсутствия латерального аппендикса). Дегенерации органа обоняния не наблюдалось. Латеральный аппендикс отсутствовал у 23 личинок.
Рассматривая результаты частичного удаления обонятельной плакоды у R. esculenta, прежде всего нужно отметить, что здесь совершенно не происходит дегенерации органа обоняния и, как следствие этого, нет случаев полного отсутствия органа. В качестве конечного результата наблюдается либо развитие нормального органа обоняния, либо образование дефектного. Количественные соотношения этих результатов представлены в табл. 7.

Таблица 7. Результаты частичного удаления обонятельной плакоды у Ranidae
Анализируя приведенные в табл. 7 данные, можно увидеть, что полученные результаты так же, как и в предыдущих случаях, отражают вполне закономерную картину количественного распределения случаев с образованием нормального и дефектного органа обоняния в зависимости от того, какая половина обонятельной плакоды была удалена. Из общего числа случаев с образованием дефектного органа обоняния - 41 на долю серий с удалением нижней и задней половин плакоды приходится 34 случая, а остальные 7 случаев относятся к серии с удалением передней половины. Аналогичные результаты были получены при соответствующих экспериментах у низших хвостатых и низших бесхвостых амфибий. Следовательно, презумптивный материал ноздри, вводного канала, хоанального канала и якобсонова органа находится у Ranidae так же, как у низших амфибий, в квадранте III. Учитывая довольно большое число случаев с образованием дефектного органа обоняния (7), относящихся к серии с удалением передней половины обонятельной плакоды, можно полагать, что этот материал захватывает частично и квадрант II (входящий в переднюю половину плакоды) в области, пограничной с квадрантом III.
Что же касается более точной локализации презумптивного материала ноздри и вводного канала, с одной стороны, и хоанального канала - с другой, то из 7 случаев дефектности органа обоняния, относящихся к серии опытов с удалением передней половины обонятельной плакоды, 4 случая обусловлены дефектностью вводного канала, что указывает на расположение его материала как в квадранте III, так и частично в квадранте II, по обе стороны разделяющей их границы. Такая локализация соответствует расположению материала ноздри и вводного канала, которое было показано в результате подобных опытов у низших хвостатых и низших бесхвостых амфибий, и поэтому в правильности ее определения не приходится сомневаться.
Более точную локализацию презумптивного материала хоанального канала по данным настоящих опытов у Ranidae непосредственно определить нельзя, так как все имеющиеся случаи отсутствия или дефектности хоанального канала сосредоточены лишь в одной серии экспериментов, а именно: в серии с удалением задней половины плакоды. Поскольку в сериях с удалением верхней и передней половин обонятельной плакоды, включающих в себя пограничные с квадрантом III квадранты IV и II соответственно, случаев с отсутствием или дефектностью хоанального канала не было, можно только с уверенностью сказать, что его материал целиком располагается в квадранте III. Однако косвенные данные, имеющие значение для более точного определения локализации материала хоанального канала у Rana, все же есть.
Во-первых, это данные по локализации рассматриваемого материала у низших хвостатых и низших бесхвостых амфибий, у которых презумптивный материал ноздри, вводного канала, хоанального канала и якоб-сонова органа располагается так же, как у Ranidae, в квадранте III, причем материал ноздри и вводного канала, с одной стороны, и хоанального канала - с другой, локализован на разных сторонах этого квадранта. Во-вторых, установлено, что локализация презумптивного материала ноздри и вводного канала у Ranidae приурочена так же, как у Hynobiidae и Discoglossidae, к стороне квадранта III, граничащей с квадрантом II. В-третьих, результаты удаления передней половины обонятельной плакоды показывают, что у Rana материал ноздри и вводного канала несколько сдвинут в сторону квадранта II и частично его захватывает. Соответственно если общий материал ноздри, вводного канала, хоанального канала и якобсонова органа занимает определенного размера участок в обонятельной плакоде, а проведенные опыты показывают некоторый его сдвиг из квадранта III в квадрант II, то находящийся на другой стороне квадранта III презумптивный материал хоанального канала должен был также сдвинуться с границы квадранта III с квадрантом IV в глубь квадранта III. При таком местоположении материала хоанального канала удаление верхней половины обонятельной плакоды не может иметь результатом отсутствие или дефектность хоанального канала, что и подтвердилось соответствующими экспериментами. Следовательно, есть все основания полагать, что презумптивный материал хоанального канала у Ranidae так же, как у Hynobiidae и Discoglossidae, локализуется около стороны квадранта III, пограничной с квадрантом IV, но только в отличие от положения этого материала у низших хвостатых и низших бесхвостых амфибий он сдвинут немного в глубь квадранта III и не выходит за его пределы.
Следует отметить, что нарушения нормального развития хоаны у высших бесхвостых амфибий менее выражены, чем у низших (4 случая из 117 возможных у Rana против 19 из 93 у Bombina) и касаются не только медиального клапана хоаны, но и латерального. Случаи неправильного распределения обонятельного эпителия главной носовой полости и случаи ее дефектности при удалении любой из половин обонятельной плакоды указывают на то, что материал главной полости у Ranidae так же, как у низших бесхвостых амфибий, либо располагается в середине плакоды, либо рассредоточен по всей плакоде.
Случаи отсутствия или дефектности латерального аппендикса относятся к 3 сериям экспериментов: удаление задней, верхней и передней половин обонятельной плакоды, причем основное количество таких случаев приходится на серии с удалением верхней и передней половин плакоды - 38 случаев из 43 имеющихся. Общим для верхней и передней половин плакоды является квадрант I, следовательно, в нем и располагается презумптивный материал латерального аппендикса. 5 случаев, имеющихся в серии с удалением задней половины плакоды, указывают на то, что этот материал захватывает частично и квадрант IV в области, граничащей с квадрантом I.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'