Хвостатые амфибии
Salamandridae. Подопытным материалом послужили два вида: Triton taeniatusn T. cristatus karelini. Были поставлены вышеописанные 4 серии опытов, сущность которых иллюстрирует прилагаемая схема операций (рис. 41). В каждой серии производилось удаление половины обонятельной плакоды, условно обозначенной: в 1-й серии - передняя половина, во 2-й - нижняя, в 3-й - задняя и в 4-й - верхняя. Благодаря такой постановке опытов можно выяснить дальнейшую судьбу четырех получившихся квадрантов, из которых I и II входят в переднюю половину плакоды, II и III - в нижнюю, III и IV - в заднюю и IV и I - в верхнюю половину (рис. 41, Б).
Из 110 оперированных эмбрионов выращено и исследовано 96, из них T. taeniatus - 74 личинки и T. cristatus karelini - 22. Разницы в результатах между обоими видами не было, и поэтому они рассматриваются вместе. По сериям опытов подопытные личинки распределились следующим образом: 1-я серия - 22, 2-я серия - 19, 3-я серия - 29 и 4-я серия - 26 личинок. Операции производились на стадиях 25 - 30 по Глезнеру, фиксация подопытных личинок - начиная со стадий 45 - 46 по Глюкзон (т. е. с момента образования хоан) и до конца метаморфоза. Препараты окрашивались по Маллори. Результаты оказались следующими.
В 1-й серии (удаление передней половины обонятельной плакоды) из 22 личинок у 9 орган обоняния отсутствовал полностью (за исключением хоаны); у 10 развился вполне нормальный орган обоняния, в 9 из них несколько уменьшенный; и только в 3 случаях образовавшийся орган обоняния оказался неполноценным. Из этих 3 случаев в двух орган обоняния представлен лишь небольшим округлым комочком клеток, безо всяких следов дифференциации на отделы, т. е. явно находится в состоянии обратного развития. В 1 случае развился орган обоняния, имеющий все отделы, кроме хоанального канала, в остальных отношениях вполне нормальный.
Во 2-й серии опытов (удаление нижней половины обонятельной плакоды) у всех 19 личинок орган обоняния имелся. В 16 случаях развился совершенно нормальный орган, в 11 из них несколько уменьшенный. В 3 случаях образовался неполноценный орган обоняния, причем в двух из них он представляет собой маленький округлый комочек клеток без дифференциации на отделы, а в 1 случае имеется орган обоняния, у которого только отсутствует ноздря и укорочен и сужен вводной канал, ведущий из ноздри в обонятельный мешок, остальные же отделы органа развиты нормально. Кроме того, в одном из случаев дегенерирующего органа обоняния хоана вместо клапана имела только небольшой выступ.
В 3-й серии опытов (удаление задней половины обонятельной плакоды), как и во 2-й, орган обоняния развился у всех 29 личинок. Нормально сформированный орган обоняния имелся только в 9 случаях, в двух из них уменьшенный. В остальных 20 случаях орган обоняния был дефектным, причем эта дефектность выражалась в отсутствии или неправильном (недостаточном) развитии следующих его частей: ноздри (отсутствовала в 12 случаях), вводного канала (отсутствовал в 8 случаях и в 4 был дефектным за счет утончения и отсутствия полости), якобсонова органа (отсутствовал в 9 случаях и в 5 был дефектным за счет уменьшения размера и отсутствия полости), хоанального канала (отсутствовал в 12 случаях и в одном был дефектным). В одном случае наблюдалось недоразвитие передней части главной полости. Кроме того, в 7 случаях хоана не имела клапана. Случаев наличия дегенерирующего органа обоняния в виде комка клеток при отсутствии дифференциации на отделы не было совсем.
В 4-й серии опытов (удаление верхней половины обонятельной плакоды) орган обоняния имелся у 21 личинки из 26, а у 5 личинок полностью отсутствовал. Только в 6 случаях из 21 развился нормально сформированный орган обоняния, в двух из них немного уменьшенный. Из остальных 15 случаев в 12 орган обоняния был дефектным, и в 3 случаях он представлял собой комок клеток, не дифференцированный на отделы. Дефектность органа обоняния была обусловлена отсутствием ноздри (7 случаев), отсутствием вводного канала (6 случаев) и его дефектностью (2 случая), отсутствием якобсонова органа (8 случаев) и его дефектностью (3 случая), отсутствием хоанального канала (12 случаев) и его дефектностью (2 случая). В одном случае отсутствовала каудальная часть главной полости. В 5 случаях хоана не имела клапана, и в одном случае он был недоразвит.
К отмечавшимся признакам дефектности органа обоняния можно было бы добавить встречающуюся довольно часто деформацию главной полости обонятельного мешка, но так как она в подавляющем большинстве случаев вызвана одновременным отсутствием ноздри и хоанального канала, что приводит к переполнению главной полости не имеющим выхода секретом, такая деформация не может быть отнесена за счет недостачи презумптивного материала этой полости и служить показателем его локализации.
Таким образом, частичное удаление обонятельной плакоды у Salamandridae имело следующие результаты.
1. Развитие вполне нормального органа обоняния, иногда несколько уменьшенного. Уменьшение размера легко объяснимо уменьшением количества исходного материала, а развитие нормального органа - регуляцией оставшегося.
2. Развитие дефектного органа обоняния, за счет отсутствия или же недоразвития какого-либо одного или нескольких его отделов: ноздри, вводного канала, хоанального канала, якобсонова органа. Результат свидетельствует о том, что в удаленной половине плакоды находился презумптивный, уже детерминированный ко времени операции, материал отсутствующего отдела.
3. Полное отсутствие органа обоняния (наличие хоаны не учитывается, так как она, как было показано выше, образуется не из материала обонятельной плакоды). Поскольку удалялась не вся обонятельная плакода, а только ее половина, то оставшаяся половина явно дегенерировала и подверглась резорбции.
4. Наличие бесформенного клеточного образования, иногда сросшегося с поверхностным эпителием или расположенного в глубине тканей, иногда сросшегося с передним мозгом, часто содержащего дегенерирующие клетки. Это явно промежуточная стадия обратного развития органа.
Количественное распределение этих результатов по сериям экспериментов показано в табл. 4.

Таблица 4. Результаты частичного удаление обонятельной плакоды у Salamandridae
Из приведенных в табл. 4 данных видно, что в количественном распределении результатов существует строгая закономерность в зависимости от того, какая половина обонятельной плакоды удалялась. Образование дефектного органа обоняния наблюдается почти исключительно (32 случая из 34 имеющихся) при удалении задней и верхней половин плакоды (при этом в 15 случаях отсутствовали одновременно все отделы органа обоняния, за исключением главной полости). При удалении как той, так и другой из этих половин всегда изымается общий для них квадрант, обозначенный на схеме номером IV (рис. 41, Б). Чистое удаление передней и нижней половин обонятельной плакоды этого квадранта не затрагивает. Ясно, что именно в квадранте IV расположен материал, из которого впоследствии образуются ноздря, вводной канал, хоанальный канал и якобсонов орган, и что этот материал ко времени произведения операций, как правило, уже детерминирован. Следовательно, в этом участке обонятельной плакоды Salamandridae находится презумптивныи материал всех основных отделов органа обоняния, за исключением главной полости.
Два случая образования дефектного органа обоняния в других сериях (один после удаления передней и один после удаления нижней половины плакоды) могут быть объяснены тем, что при операциях была захвачена также часть квадранта IV, в первом случае граничащая с квадрантом I, а во втором - с квадрантом III. Другой причиной образования дефектного органа обоняния в этих сериях экспериментов может быть то, что презумптивныи материал указанных отделов органа не лежит точно в пределах квадранта IV, а немного захватывает и прилегающие к нему участки квадрантов I и III. Чрезвычайно интересно, что в случае, имеющемся при удалении нижней половины обонятельной плакоды, у развившегося органа обоняния отсутствует ноздря и дефектен вводной канал, а в другом - при удалении передней половины плакоды, ноздря и вводной канал нормальны, но зато отсутствует хоанальный канал. Соответственно при удалении задней половины плакоды было 12 случаев отсутствия ноздри, а при удалении верхней половины - только 7. Все это дает нам более точные указания на локализацию материала этих отделов в пределах квадранта IV: материал ноздри граничит с квадрантом III и, вероятно, немного его захватывает, а материал хоанального канала - с квадрантом I, также, возможно, частично его захватывая.
Остальные три квадранта обонятельной плакоды содержат индифферентный (к моменту операций еще не детерминированный) клеточный материал, идущий на построение главной полости, т. е. самого обонятельного мешка. 2 случая дефектности главной полости (1 после удаления задней и 1 после удаления верхней половины плакоды) говорят о том, что, во-первых, этот материал в отдельных случаях может быть детерминированным и на более ранних стадиях развития и, во-вторых, о том, что он, возможно, захватывает всю срединную часть обонятельной плакоды, включая и внутренний участок квадранта IV. Эти результаты в основном согласуются с данными Тейхмана (Teichmann, 1962), который пришел к выводу, что удаление передней половины обонятельной плакоды у Triturus vulgaris и T. alpestris не влияет на формирование органа обоняния, а удаление задней половины - вызывает образование дефектов. Тейхман не мог выявить более точно локализацию материала, так как удалял только две противоположные половины плакоды. Однако в индифферентном материале I, II и III квадрантов, по-видимому, есть участок, удаление которого приводит к нарушению общего хода развития органа обоняния и в результате - либо к полному исчезновению этого органа, либо к его недоразвитию и дегенерации. Оба эти результата, вероятно, являются различными стадиями одного и того же процесса полной резорбции. Факт этот, очень интересный с точки зрения изучения механизмов развития, заслуживает отдельного исследования. К задачам настоящей работы он не имеет прямого отношения, и здесь придется ограничиться только установлением примерной локализации участка, удаление которого, по-видимому, вызывает нарушение общего хода развития органа обоняния. Квадранты III и IV явно исключаются, так как после удаления задней половины обонятельной плакоды не было ни одного случая ни отсутствия, ни обратного развития органа обоняния. Наибольшее число таких случаев (см. табл. 4) приходится на серии с удалением верхней и передней половин обонятельной плакоды. В любой из этих серий удалялся квадрант I, в нем и должен находиться этот участок, возможно захватывающий немного и квадрант II, так как при удалении нижней половины плакоды все же было 2 случая наличия дегенерирующего органа обоняния. Другой причиной резорбции органа может быть недостаток определенного объема материала, нужного для построения главной полости. Известно, что достижение определенного объема зачатка необходимо для начала процесса дифференцировки (Филатов, 1931), и вполне вероятно, что отсутствие этого фактора, вызванное экспериментально, может стать причиной обратного развития органа.
Наличие случаев с образованием нормально развитого органа обоняния в сериях опытов с удалением задней и верхней половин обонятельной плакоды, включающих в себя участок с презумптивным материалом ноздри, вводного канала, хоанального канала и якобсонова органа, говорит о том, что ко времени операции этот материал не всегда оказывался детерминированным. Развитие же уменьшенного органа обоняния объясняется уменьшением количества исходного материала.
Необходимо также отметить, что все случаи отсутствия и недоразвития хоанального клапана наблюдаются только при одновременном отсутствии хоанального канала и, следовательно, от его наличия в какой-то мере зависит правильное развитие клапана.
Hynobiidae. Эксперименты производились на эмбрионах Hynobius keyserlingii размером от 4.5 до 8.2 мм, т. е. начиная со стадии средней хвостовой почки и кончая стадией появления зачатков балансеров и жаберных бугорков. Всего было прооперировано 192 эмбриона, из них выращен, зафиксирован и исследован микроскопически 141. Фиксация проводилась на разных стадиях развития, но обязательно после срока образования хоан и якобсонова органа вплоть до конца метаморфоза. Препараты окрашивались по Маллори. В 1-й серии с удалением передней половины обонятельной плакоды насчитывается 35 экз., во 2-й серии с удалением нижней половины плакоды - 38, в 3-й серии с удалением задней половины - 35 и в 4-й серии с удалением верхней половины - 33. Результаты исследования оказались следующими.
В 1-й серии (удаление передней половины обонятельной плакоды) из 35 подопытных личинок нормальный орган обоняния развился в 32 случаях, в 23 из них уменьшенный. В 2 случаях орган обоняния оказался дефектным за счет дефектности вводного канала, в одном из этих случаев были дефектны также якобсонов орган и передняя часть главной полости, в другом было нарушено распределение обонятельного эпителия в главной полости обонятельного мешка. В третьем случае орган обоняния находился в состоянии дегенерации.
Во 2-й серии опытов (удаление нижней половины обонятельной плакоды) из 38 оперированных личинок только у 2 орган обоняния оказался нормально развитым, из них у одной он был уменьшен. Остальные 36 личинок имели дефектный орган обоняния, причем хоанальный канал был затронут во всех 36 случаях, из них в 35 он отсутствовал полностью, а в одном случае был представлен тонким клеточным тяжиком, не доходящим до хоаны. В 33 случаях дефектности органа обоняния к отсутствию хоанального канала присоединялось отсутствие или дефектность ноздри и вводного канала. Ноздря отсутствовала в 21 случае и в 9 была дефектной, вводной канал отсутствовал в 22 случаях и в 11 был дефектным. Во всех 36 случаях был затронут и якобсонов орган, из них в 27 он отсутствовал полностью, а в 9 оказался дефектным. При одновременном отсутствии ноздри и хоанального канала лишенный всякой связи с внешней средой обонятельный мешок раздувается за счет переполнения секретом и утрачивает при этом присущую ему форму. Пример дефектного органа обоняния дан на рис. 42.
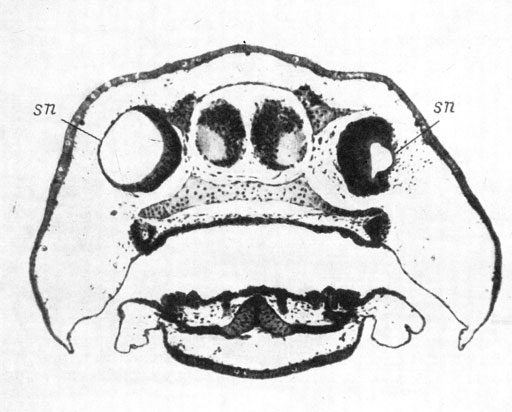
Рис. 42. Поперечный срез головы личинки Hynobius keyserlingii длиной 20 мм. Удаление нижней половины обонятельной плакоды, оперированная сторона слева. (Увел. 40)
В 3-й серии экспериментов (удаление задней половины обонятельной плакоды) из 35 подопытных личинок нормальный орган обоняния образовался у 10, у одной из них он был уменьшен. В 25 случаях развился дефектный орган обоняния. Случаев с дегенерирующим органом обоняния в этой серии, как и в предыдущей, не было. Дефектность обонятельного органа во всех 25 случаях осуществилась за счет отсутствия хоанального канала - 24 случая и его дефектности - 1 случай. Кроме того, в 18 из этих случаев к отсутствию хоанального канала присоединилось отсутствие или дефектность ноздри и вводного канала, а в 19 случаях - отсутствие якобсонова органа и в одном случае - его дефектность. Ноздря отсутствовала в 16 случаях и в одном была дефектной, вводной канал отсутствовал в 16 случаях и в двух был дефектным. В 1 случае оказалась дефектной каудальная половина главной полости. Кроме того, в 3 случаях хоана не имела клапана, и в одном случае он был очень сильно уменьшен; все эти случаи относятся к началу метаморфоза. Во всех случаях дефектности хоаны за счет клапана хоанальный канал отсутствовал.
В 4-й серии (удаление верхней половины обонятельной плакоды) из 33 подопытных личинок у 31 возник нормальный орган обоняния, из них у 26 он был уменьшен. В 1 случае орган обоняния был дефектным за счет отсутствия хоанального канала и неправильного распределения обонятельного эпителия, и в одном орган обоняния находился в дегенерирующем состоянии. Случаев отсутствия или дефектности ноздри, вводного канала и якобсонова органа в этой серии опытов не было вообще.
Таким образом, удаление различных половин обонятельной плакоды имело у Hynobiidae следующие результаты.
1. Образование нормального органа обоняния, в некоторых случаях уменьшенного.
2. Образование дефектного органа обоняния за счет отсутствия или недоразвития какого-либо одного или нескольких его отделов: ноздри, вводного канала, хоанального канала, якобсонова органа.
3. Наличие дегенерирующего органа обоняния.
Бросается в глаза, что у Hynobiidae не было ни одного случая полного отсутствия органа обоняния, как это имело место в результате подобных экспериментов у высших хвостатых амфибий сем. Salamandridae. Соответственно значительно слабее выражена у них и дегенерация органа обоняния: всего 2 случая из 141 по сравнению с 7 случаями из 96 у Salamandridae. Привожу табл. 5 с соответствующим количественным распределением результатов.
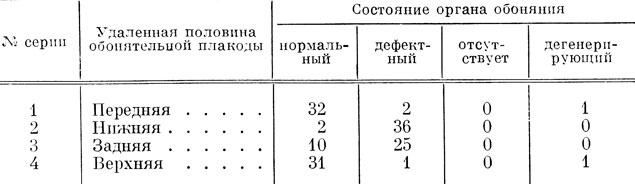
Таблица 5. Результаты частичного удаления обонятельной плакоды у Hynobiidae
Анализ данных, приведенных в табл. 5, показывает, что полученные у Hynobiidae результаты удаления различных половин обонятельной плакоды также подчиняются строгой закономерности, но закономерность эта иная, чем у высших Urodela. Основное количество случаев с образованием дефектного органа обоняния (61 случай из 64 имеющихся) падает на серии с удалением нижней и задней половин обонятельной плакоды, а не на серии с удалением задней и верхней половин плакоды, как это имело место у Salamandridae. При удалении же нижней и задней половин обонятельной плакоды общим является квадрант III, а не квадрант IV, составляющий общую часть задней и верхней половин плакоды (рис. 41, Б). Следовательно, у низших хвостатых амфибий презумптивный материал ноздри, вводного канала, хоанального канала и якобсонова органа располагается так же, как и у высших Urodela, вместе, но не в квадранте IV, как у них, а в квадранте III обонятельной плакоды.
3 случая наличия дефектного органа обоняния, падающие не на серии с удалением нижней и задней половин обонятельной плакоды, а на серии с удалением верхней и передней половин, можно объяснить так же, как в опытах над высшими хвостатыми амфибиями, излишним захватом материала или же его распространением за границы квадранта III, и точно так же эти случаи позволяют установить более четко локализацию материала ноздри и вводного канала, с одной стороны, и локализацию материала хоанального канала - с другой. В самом деле, 2 случая дефектности вводного канала отмечены при экстирпации передней половины обонятельной плакоды, следовательно, его презумптивный материал находится в квадранте III на границе с передней половиной, т. е. на границе с квадрантом II, возможно, немного его захватывая. С другой стороны, один случай отсутствия хоанального канала относится к серии с удалением верхней половины плакоды, и, следовательно, его презумптивный материал располагается в квадранте III на границе его с верхней половиной, т. е. на границе с квадрантом IV. Отсюда следует, что материал, относящийся к хоане, локализован на одной стороне квадранта, а материал вводного канала и ноздри - на другой. Эти соотношения соответствуют соотношениям в локализации материала ноздри и хоанального канала у высших хвостатых амфибий (стр. 79).
Остальные три квадранта обонятельной плакоды содержат у Hynobiidae так же, как у Salamandridae, индифферентный, по-видимому, еще не детерминированный, клеточный материал, идущий на построение главной полости обонятельного мешка, поэтому при удалении любой половины обонятельной плакоды главная полость образуется, так как материал, из которого она может развиться, занимает больше половины всей плакоды. Имеющиеся 2 случая дефектности главной полости и 2 случая неправильного распределения в ней обонятельного эпителия говорят о возможности сдвига детерминации материала на более ранние сроки.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'