Бесхвостые амфибии
Если у хвостатых амфибий развитие якобсонова органа протекает одинаково у всех исследованных видов (отмечается только некоторая разница в ориентировке закладки), то у бесхвостых амфибий можно выделить три различных типа развития якобсонова органа, которые отличаются друг от друга как по самому ходу развития, так и по окончательным результатам, - тип Xenopus, тип Bombina и тип Rana. Поэтому нецелесообразно давать общее сравнительное описание хода развития якобсонова органа у бесхвостых амфибий, как это было сделано в отношении хвостатых, а необходимо рассмотреть все три типа развития в отдельности.
У Xenopus laevis якобсонов орган возникает очень рано, показывая в этом отношении большое сходство с закладкой его у амниот (Foske, 1934). Он обосабливается в виде округлого утолщения на передне-вентральном участке органа обоняния и направлен в момент своего возникновения вентро-медиально. В это время обонятельный орган представляет собой только еще очень мелкую обонятельную ямку (стадия 32 по Ньюкопу и Фаберу). На стадии 33/34 обонятельная ямка немного углубляется, и ее полость начинает заходить в глубь шарообразного выступа якобсонова органа. Все это происходит еще до начала образования хоанального выроста обонятельной ямки, который возникает лишь на стадии 35/36 (рис. 15, А). В процессе дальнейшего развития происходит углубление обонятельной ямки и превращение ее в мешок, в результате чего якобсонов орган оказывается направленным строго медиально (рис. 15, В, стадия 39). По мере последующего развития и все большего уплощения головы личинки обонятельный мешок растет назад, а якобсонов орган остается на месте и, по-прежнему отходя от области, близкой к ноздре, располагается под ней и впереди от обонятельного мешка.
На стадии 45 задняя стенка якобсонова органа образует небольшой вырост (закладка якобсоновой железы), который затем разрастается назад, вдоль медиальной стенки обонятельного мешка (стадии 46 - 50). Сам якобсонов орган растет вперед, и его передний слепой конец лежит намного впереди ноздри (стадия 53). Якобсонова железа сильно разрастается, она окружает задний конец якобсонова органа и заходит под вентральную стенку обонятельного мешка (стадия 54).
Начиная с предметаморфозной стадии 56 и во время метаморфоза происходит усложнение обонятельного мешка путем образования добавочных полостей, подробно описанное Феске. Одновременно с этим процессом хоана, которая на протяжении всего предыдущего развития находилась непосредственно под ноздрей, быстро смещается каудально. В результате этого смещения главная полость обонятельного мешка, ранее располагавшаяся кзади от ноздри и хоаны, лежавших на одной вертикальной плоскости, передвигается вперед, а якобсонов орган - назад, под нее, и смещается несколько латерально, благодаря чему он к середине метаморфоза лежит не медиально от обонятельного мешка, а вентрально от него, причем образует выступ, немного заходящий на латеральную сторону (стадия 59, рис. 30). Это расположение, сохраняемое и дефинитивно, и дало Феске возможность говорить о промежуточном состоянии (между характерными для хвостатых и для бесхвостых амфибий) якобсонова органа у Xenopus. Следует отметить, что латеральный выступ якобсонова органа Xenopus является закладкой и на последующих стадиях началом заднего отрезка бокового носового желобка, переходящего далее кзади в небольшую нёбную борозду, продолжающуюся до середины хоаны. По мере своего развития якобсонов орган Xenopus отстает в росте от обонятельного мешка и относительно сильно уменьшается в размере.
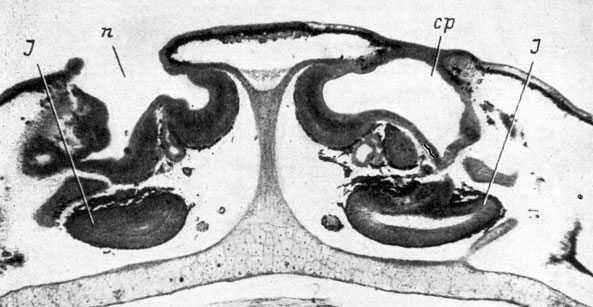
Рис. 30. Поперечный срез через обонятельную область метаморфизмрующей личинки Xenopus laevis длиной 55 мм, стадия 59. (Увел. 40)
Развитие якобсонова органа у обыкновенной жерлянки - Bombina bombina - начинается с образования небольшого утолщения обонятельного эпителия в вентро-медиальном углу обонятельного мешка, на границе с индифферентным эпителием, выстилающим вентро-латеральную стенку мешка (личинка длиной 9 мм, рис. 31, А), что представляет поразительное сходство с закладкой якобсонова органа у хвостатых амфибий (рис. 28, А). К этому времени хоанальный канал и хоана уже образованы (см. стр. 32). Полость обонятельного мешка начинает заходить в закладку якобсонова органа не сразу, а немного позже, при этом она направлена вентро-медиально (личинка длиной 9.5 мм). Затем закладка якобсонова органа вытягивается в вырост, отходящий в это время уже не от вентро-медиального угла обонятельного мешка, а от медиальной части его вентральной стенки, причем направлен он почти вентрально (личинка длиной 11 мм). При последующем развитии направление якобсонова органа становится строго вентральным, так же ориентирована и его полость. Место отхождения якобсонова органа от обонятельного мешка при этом перемещается на середину его вентральной стенки (личинка длиной 15 мм). Одновременно якобсонов орган начинает разрастаться вперед под обонятельным мешком, и медиальная стенка его переднего конца образует шарообразный выступ с полостью - закладку якобсоновой железы, направленную медиально. В процессе дальнейшего развития место отхождения якобсонова органа от обонятельного мешка сдвигается на латеральную половину его вентральной стенки, а разросшийся передний конец якобсонова органа ориентируется в латеральную сторону.
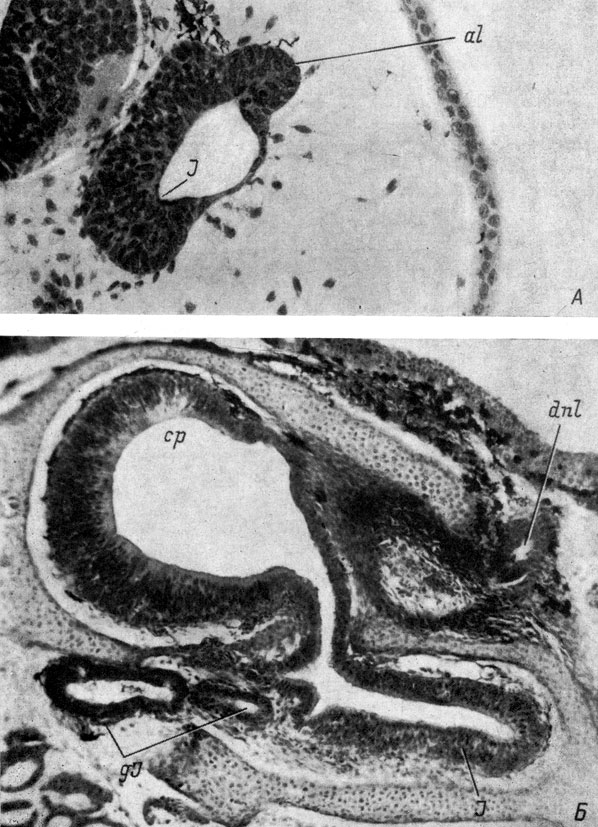
Рис. 31. Поперечные срезы через правый орган обоняния личинок Bombina bombina длиной 9 мм (А, увел. 170) и 50 мм (Б, увел. 80)
Одновременно со смещением места отхождения якобсонова органа из вентро-медиального угла обонятельного мешка в вентро-латеральный угол, граничащий с этим местом индифферентный эпителий, ранее составлявший вентральную стенку и вентро-латеральный угол мешка, также сдвигается латерально, а затем дорсально, образуя в конечном результате латеральную стенку обонятельного мешка (рис. 31, Б). Это одновременное смещение соседних, отличающихся друг от друга, участков стенки обонятельного мешка в одном направлении явно показывает, что причиной этих процессов является поворот мешка вокруг своей длинной оси. Дополнительным доказательством такого поворота служит тот факт, что к моменту достижения местом отхождения якобсонова органа вентро-латерального угла обонятельного мешка (личинка длиной 39.5 мм) передний конец органа лежит еще под медиальной половиной мешка, т. е. в своем передвижении он отстает от заднего конца, связанного со стенкой мешка и поэтому смещающегося с большей скоростью. Такое отставание в передвижении создает временную медиальную ориентировку якобсонова органа (личинка длиной 42 мм).
В предметаморфозный период начинается дифференцировка обонятельного мешка и образование добавочных полостей. Между главной носовой полостью, в которую превращается большая часть обонятельного мешка и местом отхождения якобсонова органа возникает выпячивание средней носовой полости (личинка длиной 42 мм), стенки которой состоят из индифферентного эпителия. Эта средняя полость сначала отходит от вентро-медиального угла передней половины главной носовой полости и имеет вид плоской складки, направленной латерально и подстилающей вентральную стенку передней половины главной полости. Якобсонов орган соединяется при этом уже не непосредственно со стенкой главной носовой полости, а с латеральным краем средней полости. Поскольку последний сильно разросся латерально, то, отходя от него, якобсонов орган в целом имеет медиальную ориентировку, хотя его задний конец располагается под латеральной половиной как средней, так и главной полости. При дальнейшем развитии средней носовой полости место ее отхождения от главной перемещается из вентро-медиального в вентро-латеральное положение, благодаря чему сдвигается еще более латерально не только место отхождения якобсонова органа от обонятельного мешка, но и сам орган, который располагается в результате латерально от главной носовой полости (личинка длиной 45 мм). Передний медиальный купол главной носовой полости, находившийся раньше сзади якобсонова органа, теперь лежит над его каудальной половиной.
Средняя носовая полость, разрастаясь, образует изгиб в медиальную сторону (личинка длиной 49 мм), а затем вторичный вентральный изгиб, на который и смещается место отхождения якобсонова органа, направленного латерально как от средней, так и от главной носовой полости (личинка длиной 50 мм, рис. 31, Б). Кзади якобсонов орган переходит в довольно плоскую складку, состоящую из индифферентного эпителия - боковой носовой желобок. Якобсонова железа располагается медиально от органа обоняния и после образования хрящевой обонятельной капсулы лежит снаружи от нее, рядом с межносовой железой (характерно только для Bombina, у всех остальных исследованных видов амфибий якобсонова железа находится внутри хрящевой капсулы). К концу метаморфоза положение якобсонова органа по своей строго латеральной ориентации практически не отличается от его положения у хвостатых амфибий (рис. 33).
У лягушек рода Rana якобсонов орган также возникает в виде выступа стенки медио-вентрального угла обонятельного мешка и при своем возникновении направлен медиально (личинка R. esculenta длиной 8 мм). Его появление совпадает с моментом возникновения хоанального канала. У личинки длиной 9.5 мм якобсонов орган хотя и располагается по-прежнему в медио-вентральном углу обонятельного мешка, но уже направлен не медиально, а почти вентрально. Немного позже якобсонов орган сдвигается полностью на вентральную стенку мешка, помещаясь в ее медиальной части (R. esculenta длиной 10.5 мм, рис. 32, А). Направление его при этом строго вентральное. В это же время якобсонов орган начинает разрастаться вперед, и в дорсо-медиальном углу его передней части появляется выступ - закладка якобсоновой железы.
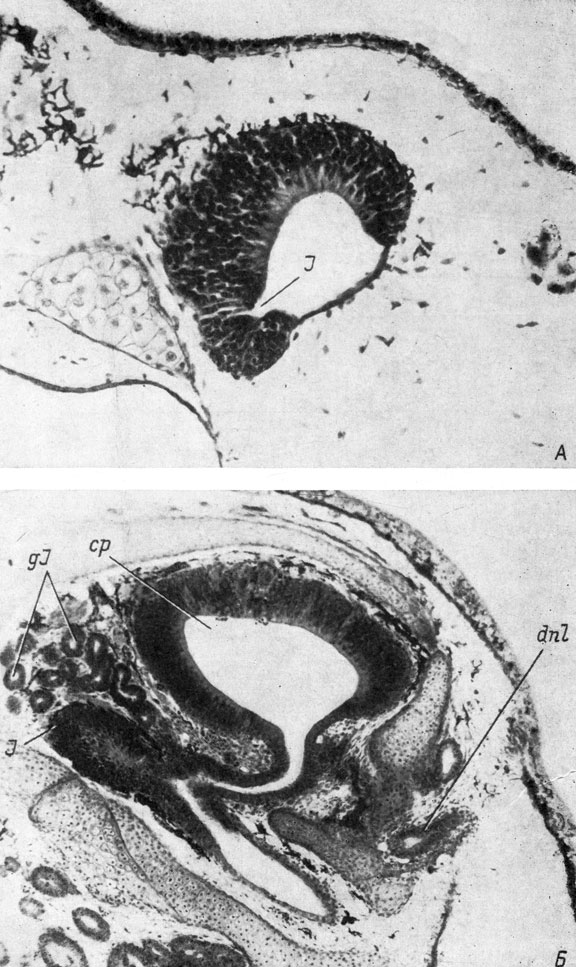
Рис. 32. Поперечные срезы через правый орган обоняния личинок Rana esculenta длиной 10.5 мм (А, увел. 170) и R. temporaria длиной 36 мм (Б, увел. 80)
Дальнейшего сдвига места отхождения якобсонова органа в латеральную сторону у Rana не наблюдается, а, наоборот, у личинки R. esculenta длиной 13 мм восстанавливаются более ранние соотношения, и якобсонов орган опять отходит от вентро-медиального угла обонятельного мешка, ориентируясь также вентро-медиально. Передний конец якобсонова органа по мере разрастания вперед превращается в складку, направленную медио-дорсально (личинка длиной 15 мм). Якобсонова железа, увеличиваясь в размере, располагается медиально и каудально от якобсонова органа. В процессе довольно длительного дальнейшего развития якобсонов орган сохраняет свое вентро-медиальное отхождение от передней части главной носовой полости, которая начинает образовывать добавочные полости только с наступлением метаморфоза.
В начале метаморфоза (личинки R. temporaria длиной 30 - 33 мм) стенка передней части главной носовой полости образует вырост, состоящий из индифферентного эпителия и направленный медиально. Этот вырост отделяет якобсонов орган от главной носовой полости и является зачатком средней полости. Латеральный край средней носовой полости разрастается в виде слепого мешка вперед, а медиальная ее часть распространяется до якобсонова органа, в результате чего место отхождения последнего перемещается на медиальный участок средней полости (личинка R. temporaria длиной 36 мм, рис. 32, Б). Вторичный латеральный изгиб средней носовой полости, одновременно являющийся продолжением латеральной части якобсонова органа, переходит далее кзади в боковой носовой желобок. Якобсонов орган, отходя теперь уже от медиальной части средней носовой полости, направлен медиально и несколько дорсально, достигая своим передним слепым концом уровня ноздри. Это положение он сохраняет и в дефинитивном состоянии.
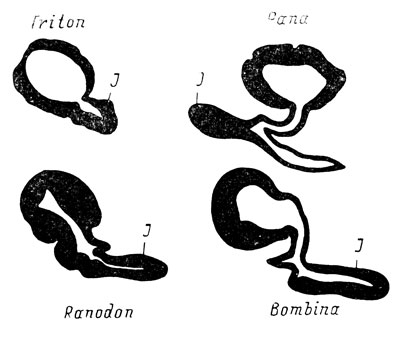
Рис. 33. Схема дефинитивного положения якобсонова органа у хвостатых и бесхвостых амфибий
Таковы основные типы развития якобсонова органа бесхвостых амфибий. Развитие его у чесночницы Pelobates fuscus в основном происходит по типу, наблюдающемуся у Rana. Некоторое расхождение наступает лишь на конечных, метаморфозных, стадиях развития, что отражается на дефинитивном положении органа. Якобсонов орган отходит так же, как и у Rana, от медиальной части средней носовой полости, но направлен не медио-дореально, а строго медиально. Его передний конец (и весь орган в целом) располагается не медиально от передней части главной носовой полости, а непосредственно под ней, находясь на метаморфозных стадиях под ее латеральной половиной. К концу метаморфоза задний отрезок якобсонова органа лежит под средней линией главной носовой полости, но его самый передний конец остается под латеральной ее половиной. Следовательно, несмотря на то что развитие якобсонова органа у P. fuscus протекает по типу, характерному для Rana, в конце развития он приобретает положение, в очень большой степени сходное с положением якобсонова органа у Xenopus.
Таким образом, различные типы развития якобсонова органа у бесхвостых амфибий приводят к диаметрально противоположному дефинитивному положению органа, начиная от медиального у Rana (и у большинства бесхвостых амфибий - Helling, 1938) и кончая латеральным у Bombina, совершенно идентичным положению у хвостатых амфибий (рис. 33). Все же, несмотря на различный окончательный результат, у всех бесхвостых амфибий прослеживается хотя бы первоначальный сдвиг медиальной закладки якобсонова органа в латеральную сторону, приводящий в случае его яркой выраженности к латеральному положению органа (тип развития Bombina), такому же, как и у хвостатых амфибий, а у большинства Anura, этот сдвиг завуалирован последующими изменениями процесса развития, который дает в конечном итоге либо чисто медиальное положение органа, либо промежуточное между медиальным и латеральным. Что же касается якобсоновой железы, то ход ее развития и ее дефинитивное положение не имеют принципиальных отличий ни среди самих бесхвостых амфибий, ни в сравнении с развитием этой железы у Urodela.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'