Хвостатые амфибии
Ход развития якобсонова органа в основном одинаков у всех хвостатых амфибий, существуют только некоторые различия в местоположении, вернее, в ориентировке, самой ранней его морфологической закладки. Такая самая ранняя распознаваемая закладка якобсонова органа представляет собой небольшое утолщение обонятельного эпителия еще не расчлененного обонятельного мешка, находящееся в его вентро-медиальном углу и располагающееся на границе с индифферентным эпителием, составляющим в это время вентральную стенку органа обоняния. У всех хвостатых амфибий закладка якобсонова органа возникает через некоторое время после образования хоан.
У Hynobiidae (личинки Hynobius keyserlingii длиной 12.5 мм и Ranodon sibiricus длиной 21 - 22 мм) утолщение стенки обонятельного мешка, представляющее собой закладку якобсонова органа, направлено в медиальную сторону, и в него с самого момента его образования начинает заходить полость обонятельного мешка, также ориентированная медиально (рис. 28, А, А′). У Triton cristatus karelini закладка находится тоже в вентро-медиальном углу обонятельного мешка, но направлена вентро-медиально (личинка длиной 10.5 мм, стадия 42 по Глюкзон). У личинок аксолотля длиной 11.5 - 12 мм (стадия 37 по Гаррисону; Воронцова и др., 1952) местоположение закладки якобсонова органа и ее ориентировка так же, как у кавказского подвида гребенчатого тритона, вентро-медиальные. У личинок обыкновенного (длина около 10 мм, стадия 46 по Глюкзон) и испанского (длина 12 мм, стадия 37 по Гальену и Дюроше) тритонов, несмотря на то что закладка якобсонова органа располагается в том же вентро-медиальном углу обонятельного мешка, она целиком относится к вентральной его стенке и ориентирована вентрально. У последних четырех видов полость якобсонова органа возникает не сразу, а на более поздних стадиях, причем у аксолотля (личинки длиной 12 - 13 мм, стадия 38) она направлена в момент образования вентро-медиально, а у личинок обыкновенного (длина 12 мм, стадия 54) и испанского (длина 15 мм, стадия 42) тритонов ориентировка полости сразу имеет вентральное направление (рис. 29, А).
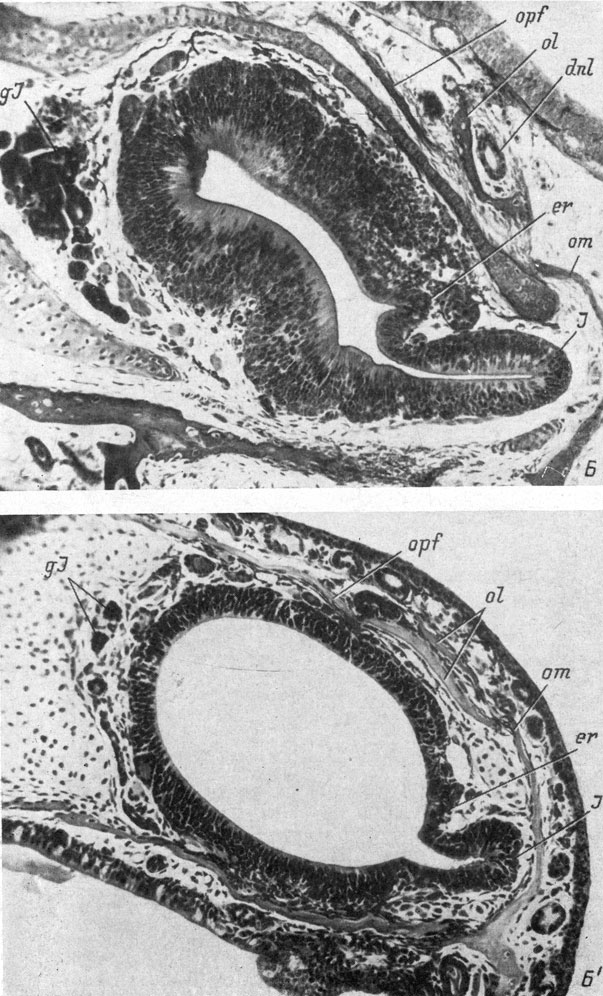
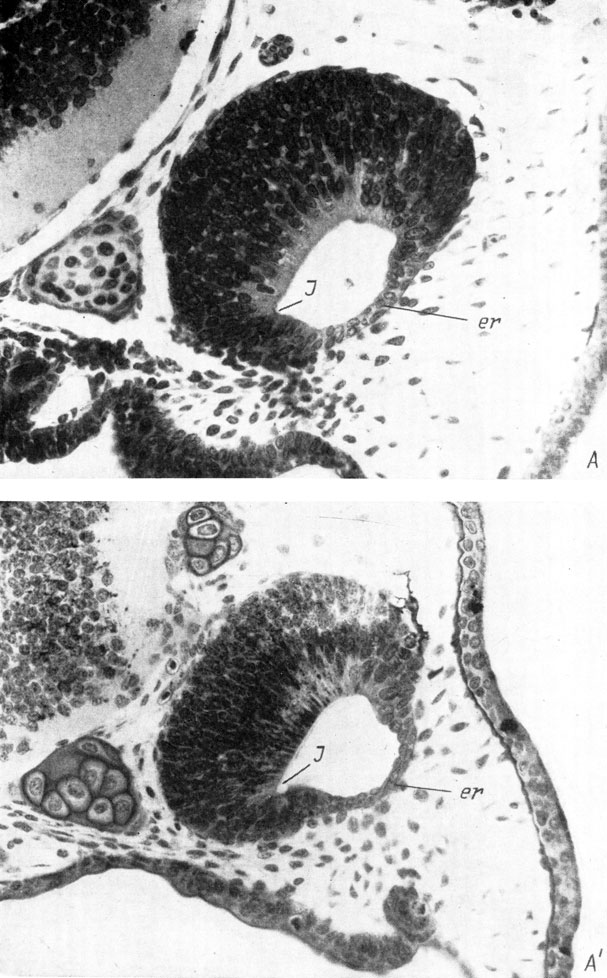
Рис. 28. Поперечные срезы через правый орган обоняния личинок Ranodon sibiricus длиной 21- 22 мм (А, увел. 170) и 74 мм (Б, увел. 90) и Hynobius keyserlingii длиной 12.5 мм (А′, увел. 170) и 29 - 30 мм (Б′, увел. 90)
В процессе дальнейшего развития якобсонова органа он постепенно изменяет место своего отхождения от обонятельного мешка и свое положение от вентро-медиального к вентральному, затем к вентро-латеральному и, наконец, занимает латеральное положение по отношению к обонятельному мешку, отходя от его вентро-латерального угла (рис. 28, Б, Б′; 29, Б). Одновременно происходит соответствующее изменение направления полости якобсонова органа на латеральное. В пользу точки зрения, по которой описываемый процесс представляет собой не смещение места отхождения якобсонова органа по вентральной стенке обонятельного мешка, а является результатом вращения самого мешка, говорит то обстоятельство, что зона индифферентного эпителия, находившаяся рядом с местом отхождения якобсонова органа и располагавшаяся в начале его развития по вентральной стенке обонятельного мешка (рис. 28, А, А′), также передвигается в латеральную сторону и в результате оказывается на латеральной стенке последнего, над якобсоновым органом (рис. 28, Б, Б′).
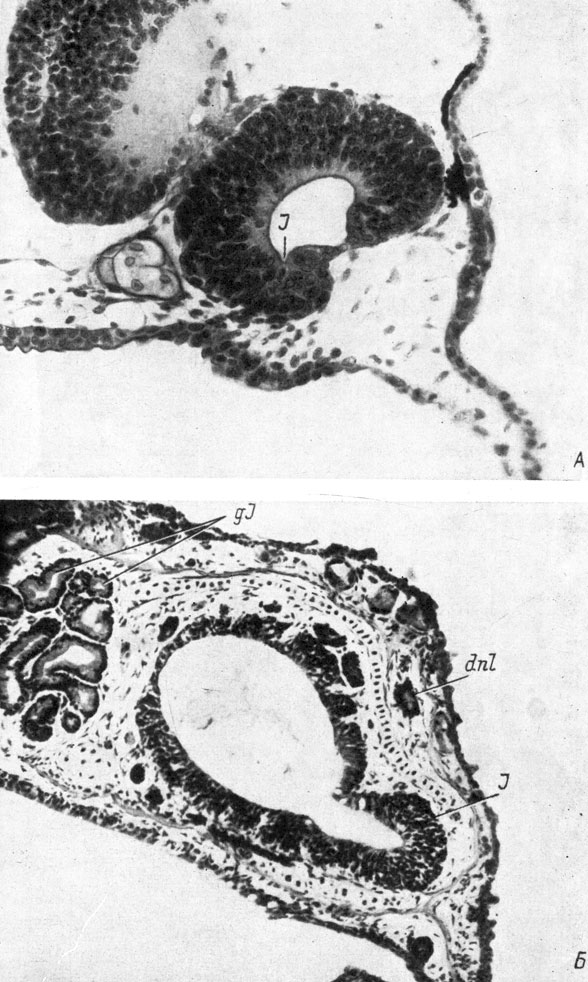
Рис. 29. Поперечные срезы через правый орган обоняния личинки Triton taeniatus длиной 12 мм (А, увел. 170) и метаморфизировавшего тритона длиной 30 мм (Б, увел. 90)
В ходе своего развития, в то время когда якобсонов орган занимает вентральное положение, он начинает разрастаться вперед и несколько назад. Медиальная стенка передней части якобсонова органа образует вырост, направленный в медиальную сторону и подстилающий вентральную стенку обонятельного мешка, - это закладка якобсоновой железы. Начало развития якобсоновой железы отмечается у обыкновенного тритона на стадии 59 (длина личинок 15 - 16.5 мм), у кавказского подвида гребенчатого тритона на стадии 52 (длина личинок 19 мм), у испанского тритона на стадии 48 (длина личинок 20.5 - 21 мм), у аксолотля на стадии 38 - 39 (длина личинок 15 - 16 мм), у Hynobius keyserlingii у личинок длиной 15 мм и у Ranodon sibiricus у личинок длиной 28 - 29.5 мм. Якобсонова железа растет в дальнейшем между медиальной стенкой обонятельного мешка и хрящевой обонятельной капсулой, основная ее масса располагается в конечном итоге на уровне верхней части обонятельного мешка с его медиальной стороны (рис. 28, Б, Б′; 29, Б). Выводной проток якобсоновой железы всегда открывается в медиальной стенке передней части якобсонова органа. Дефинитивный якобсонов орган у всех хвостатых амфибий лежит латерально от обонятельного мешка и несколько ниже его, соединяясь с последним в латеро-вентральном углу его срединной части.
Таким образом, ход развития якобсонова органа весьма сходен у всех хвостатых амфибий. У более примитивных Urodela закладка якобсонова органа представляет собой медиальное выпячивание стенки обонятельного мешка так же, как и у остальных наземных позвоночных. То, что у более высоко организованных хвостатых амфибий закладка якобсонова органа имеет не медиальную, а вентральную ориентировку, говорит только о выпадении у высших Urodela ранних стадий развития органа (довольно частое явление в развитии признаков более высоко организованных форм). Доказательством тому служит промежуточное между обоими положениями закладок (медиальным у Hynobiidae и вентральным у Triton taeniatus и Pleurodeles waltlii) состояние их у кавказского подвида гребенчатого тритона и у аксолотля. Следовательно, исходным для хвостатых амфибий является медиальное положение закладки якобсонова органа, а его латеральное дефинитивное положение вторично.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'