Развитие собственно хоан и хоанального канала хвостатых амфибий
Раннее развитие органа обоняния протекает у различных видов хвостатых амфибий весьма сходно, поэтому отпадает необходимость отдельного описания процесса развития для каждого из исследованных видов и вполне можно ограничиться сравнительным изложением данных, останавливаясь на расхождениях в тех случаях, когда они имеются. Основу описания составит процесс развития собственно хоан и хоанального канала у Ranodon sibiricus и Hynobius keyserlingii поскольку эти виды являются наиболее примитивными, а у первого из них развитие органа обоняния ранее вообще не исследовал ось и изучено впервые автором (Медведева, 1961а, 1964). К сожалению, для H. keyserlingii и R. sibiricus нет таблиц нормального развития, и поэтому по ходу изложения в скобках указываются примерно соответствующие приводимым стадии развития обыкновенного тритона по Глезнеру (до стадии 41) и по Глюкзон для более поздних стадий (Воронцова и др., 1952) и испанского тритона по Гальену и Дюроше (Gallien et Durocher, 1957).
На стадии ранней хвостовой почки обонятельные плакоды R. sibiricus и других видов хвостатых амфибий (стадия 21 - 22 у обыкновенного и у испанского тритонов) представляют собой небольшие утолщения чувствующего слоя эктодермы и располагаются в самой ростральной части головы, на ее вентро-латеральных сторонах. В процессе последующего развития плакода превращается в массивное, хорошо отграниченное от покровов утолщение, хотя края ее по-прежнему полого переходят в окружающую покровную эктодерму. На стадии средней хвостовой почки (стадии 23 - 25 у обыкновенного и у испанского тритонов) на поперечных срезах видно, что плакода почти полностью заполняет треугольное пространство между передним мозгом и глазным пузырем. Сагиттальные срезы показывают, что обонятельная плакода лежит несколько впереди и немного вентральнее медиальной половины глазного пузыря и находится на довольно значительном расстоянии от энтодермальной передней кишки (рис. 21). Ротового впячивания еще нет, но кишка своим передним концом подходит очень близко, в медианном участке почти вплотную, к покровной эктодерме и переднему мозгу. Боковые участки переднего конца кишки находятся несколько дальше от эктодермы, чем медианный ее участок, и пространство между покровами и передней кишкой оказывается заполненным мезенхимой.
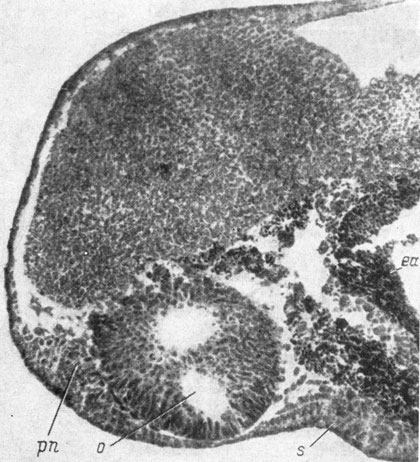
Рис. 21. Сагиттальный срез головы эмбриона Hynobius keyserlingii длиной 4.5 мм. (Увел. 80)
Одновременно с возникновением в средней части обонятельной плакоды углубления, являющегося зачатком ноздревого отверстия, начинается образование ротового впячивания. В отличие от бесхвостых амфибий у Urodela ротовое впячивание эктодермального материала не влечет за собой одновременного образования углубления или ямки, а ограничивается врастанием идущих навстречу друг другу с ростральной и каудальной сторон слоев эктодермы. В латеральных участках ротового впячивания эти слои эктодермы плотно прилегают друг к другу и, по всей видимости, срастаются в момент встречи, а в медианной части впячивания они сразу же начинают обрастать подходящую здесь вплотную к покровам энтодерму переднего конца кишки опять-таки с ростральной и каудальной сторон. Именно эта особенность процесса образования ротового впячивания у хвостатых амфибий, состоящая в том, что у них не возникает никакого ротового углубления, обусловливает и отсутствие у них выраженной рото-глоточной мембраны, определяющей границу между эктодермой и энтодермой в передней кишке.
Наружная область ротового впячивания представляет в это время поперечную, более широкую в своей медианной части, полосу, немного заходящую на боковые стороны головы. Поскольку в своих латеральных участках энтодермальная передняя кишка не подходит так близко к покровной эктодерме, как в медианном, то на этой стадии развития (стадия 25 - 26 у обыкновенного и у испанского тритонов) врастающие внутрь боковые края ротового впячивания, включая участки, находящиеся в одной сагиттальной плоскости с обонятельными плакодами, еще не соединяются с энтодермой передней кишки и остаются чисто эктодермальными (рис. 22, А). Но в медианной части ротового впячивания врастающая эктодерма сразу же приходит в соприкосновение с лежащей вплотную к ней ростральной частью энтодермальной передней кишки и начинает обрастать последнюю с ростральной и каудальной сторон (самый медианный участок врастающей с ростральной стороны эктодермы образует гипофизарную закладку - карман Ратке). В результате в медианной части ротового впячивания (или, точнее, врастания) с момента начала его образования наступает соединение с энтодермальной передней кишкой, и последняя таким образом становится с самого начала этого процесса экто-энтодермальной (рис. 22, Б). Поэтому хотя на этих стадиях развития эктодерму очень легко отличить от энтодермы, так как клетки последней сплошь забиты желточными гранулами, о границе между эктодермой и энтодермой в общепринятом смысле слова говорить не приходится, поскольку передняя кишка, т. е. будущая ротовая полость, оказывается состоящей из двух зародышевых листков, из которых эктодерма располагается в наружных слоях кишки, а энтодерма - во внутренних, причем последняя доходит в своем переднем участке до самой поверхности тела эмбриона.
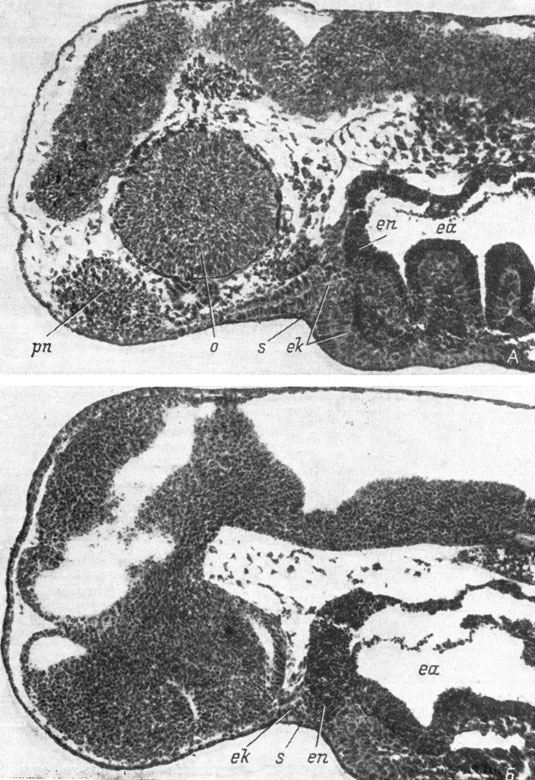
Рис. 22. Сагиттальные срезы (латеральный - А и медианный - Б) головы эмбриона Hynobius keyserlingii длиной 6 мм. (Увел. 76)
Несколько позже (стадия 27 - 28 у обыкновенного и у испанского тритонов), одновременно с образованием глубокой обонятельной ямки, этот процесс обрастания рострального конца передней кишки эктодермой захватывает и боковые участки кишки и ротового впячивания. Таким образом, вся передняя кишка оказывается одетой снаружи эктодермой, а внутри состоящей из энтодермы. Соединение ротового впячивания с передней кишкой протекает совершенно одинаково у всех исследованных видов хвостатых амфибий. Фотографии сагиттальных срезов головы личинок Hynobius keyserlingii (рис. 23, А) и Necturas maculatus (рис. 23, Б) показывают соотношение эктодермы и энтодермы в боковых частях передней кишки, лежащих в тех же сагиттальных плоскостях, которые проходят через обонятельные ямки. На срезе головы N. maculatus, кроме того, очень хорошо видно, что утолщенный эпителиальный слой, находящийся между обонятельной ямкой и ротовым впячиванием, состоит из закладки инфраорбитальной плакоды органов боковой линии (ср. с рис. 19, А).
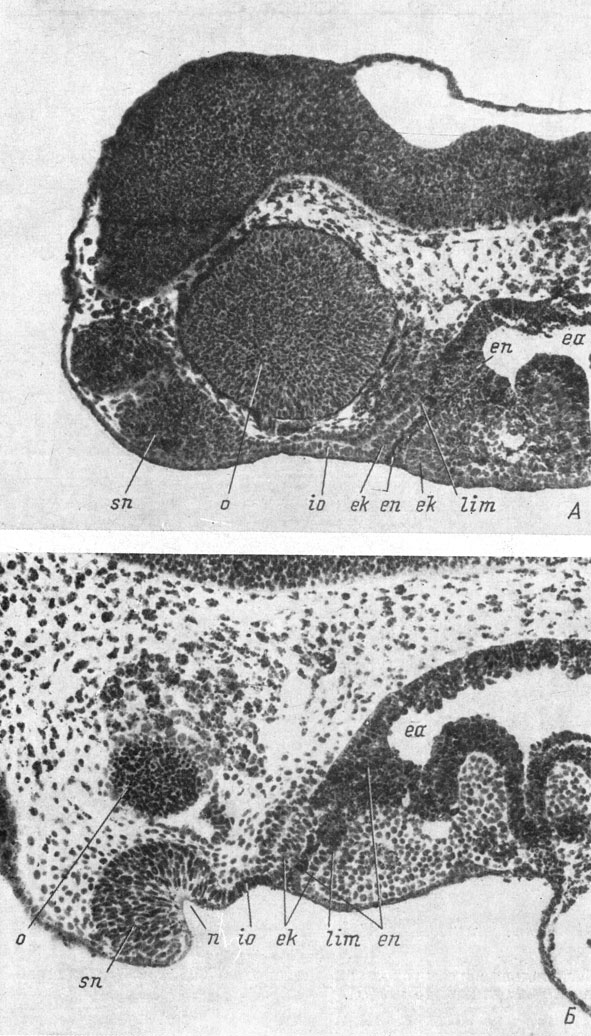
Рис. 23. Сагиттальные срезы головы эмбрионов Hynobius keyserlingii длиной 7.5 мм (А) и Necturus maculatus длиной 12 мм (Б). (Увел. 76)
Расстояние между обонятельной ямкой и передней кишкой, а также соответственно между первой и ротовым впячиванием по-прежнему остается довольно значительным, особенно у Ranodon sibiricus и Hynobius keyserlingii. Однако по мере дальнейшего углубления обонятельной ямки и превращения ее в мешок начинается постепенный сдвиг наружного носового отверстия по вентральной поверхности морды в каудальную сторону, по направлению к ротовому впячиванию. При этом обонятельная ямка, лежавшая вначале на самом переднем конце головы и впереди глаза (рис. 21), оказывается на ее вентральной поверхности, под глазом. Поскольку наиболее интенсивно перемещается наружная часть обонятельного мешка, то наружное носовое отверстие, которое на более ранних стадиях развития было направлено вентрально, теперь оказывается направленным в каудальную сторону, а несколько отстающая при этом передвижении внутренняя часть мешка, образующая его дно, лежит теперь впереди остальной части обонятельного мешка и ноздри и направлена в сторону переднего конца морды (рис. 24, А, Б). Такое положение обонятельного мешка с очевидностью свидетельствует о перемещении его наружной части вместе с ноздрей назад, по направлению к ротовому впячиванию, причем ведущую роль в этом процессе явно играет передвижение ноздри, влекущей за собой наружную часть обонятельного мешка.
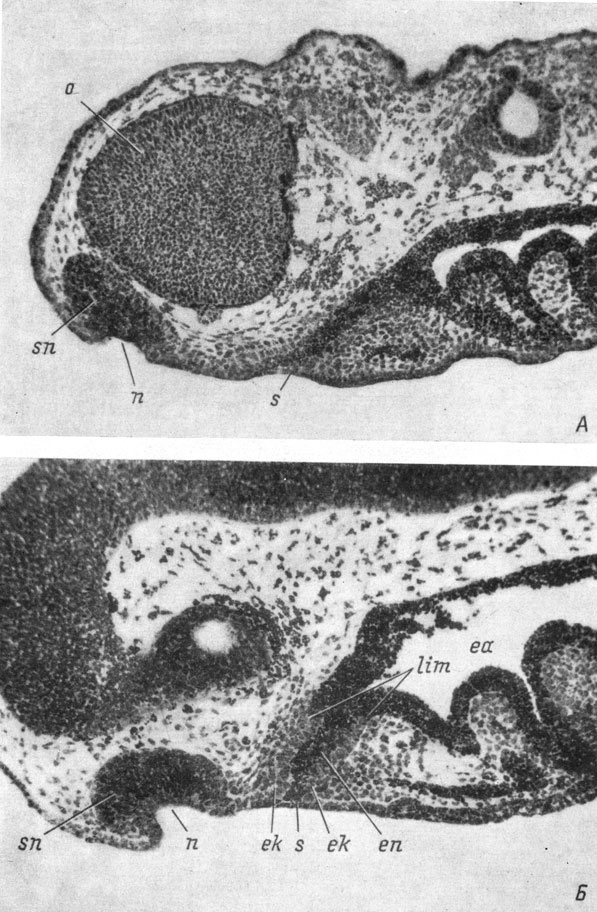
Рис. 24. Сагиттальные срезы головы эмбрионов Hynubius keyserlingii длиной 8.5 мм (А) и Ranodon sibiricus длиной 13 мм (Б). (Увел. 76)
Одновременно, по-видимому, имеет место некоторое смещение ротового впячивания и конца передней кишки в ростральную сторону, по направлению к обонятельному мешку, так как ротовое впячивание находится теперь не каудальнее глаза, как раньше, а под ним. В результате этих перемещений к моменту начала образования хоанального канала (стадии 33 - 35 у обыкновенного тритона, стадия 32 у испанского тритона) обонятельный мешок и передняя кишка оказываются лежащими очень близко друг от друга, а расстояние от каудальной поверхности мешка до ростральной стороны будущей ротовой полости - очень маленьким. Описываемый процесс смещения наружных носовых отверстий назад, а будущей ротовой полости - вперед гораздо явственнее выражен у Ranodon sibiricus и Hynobius keyserlingii, чем у остальных исследованных видов, ввиду того что у последних обонятельные плакоды закладываются несколько каудальнее переднего конца головы, а ротовое впячивание начинает образовываться ростральнее и ближе к обонятельным ямкам, чем у представителей сем. Hynobiidae. Поэтому у более высоко организованных хвостатых амфибий этот процесс значительно сглажен и не так бросается в глаза.
В это же время в боковых участках передней кишки, а именно в тех, которые ближе всего подходят к обонятельным мешкам, образуются симметрично расположенные выросты, направленные рострально (рис. 25, А). Возникновение и развитие этих выростов происходит до появления каких-либо признаков образования хоанального выроста со стороны обонятельного мешка (рис. 25, Б). Поскольку передняя кишка со своей наружной стороны одета эктодермой, а внутри заполнена желточной энтодермой, то каждый из образующихся выростов кишки оказывается состоящим в основном из эктодермальных клеток, но во внутреннюю его часть заходит из кишки желточная энтодерма (рис. 25, В). Вырост растет вперед к обонятельному мешку соответствующей стороны.
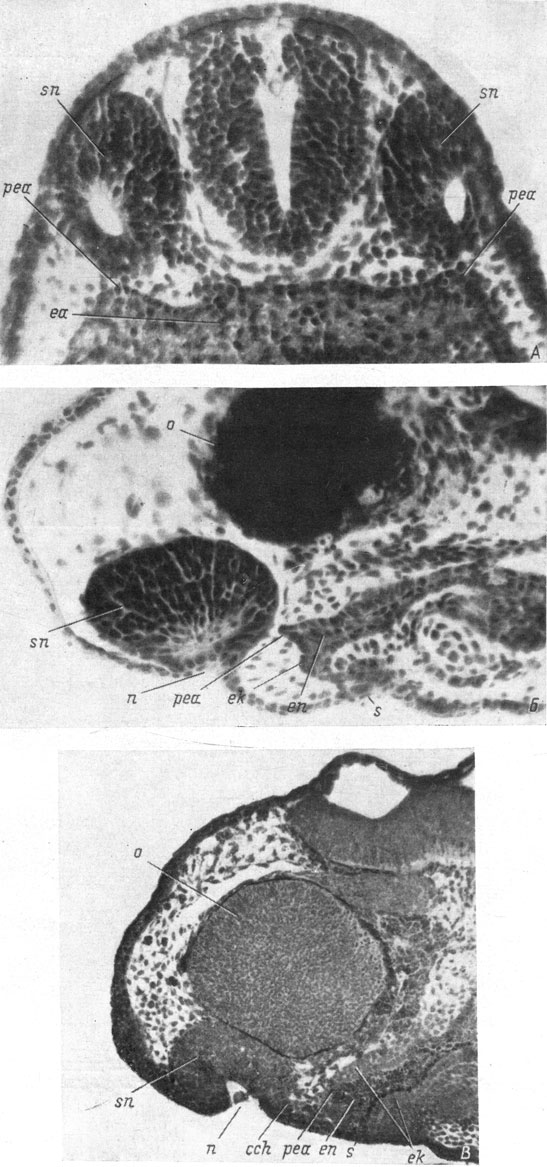
Рис. 25. Фронтальный срез головы эмбриона Triton taeniatus длиной 7.5 мм (А, увел. 160) и сагиттальные срезы головы эмбрионов Т. taeniatus длиной 8 мм (Б, увел. 160) и Ranodon sibiricus длиной 14 мм (В, увел. 76)
Образование хоанального выроста обонятельного мешка начинается (стадии 36 - 38 у обыкновенного тритона, стадия 33 у испанского тритона) на самой границе обонятельного и индифферентного эпителия, у каудального края ноздри, когда вырост передней кишки уже имеется. Рост хоанального выроста обонятельного мешка осуществляется за счет интенсивного размножения клеток переходной зоны между обонятельным и индифферентным эпителием. С начала образования хоанального выроста в него начинает заходить полость обонятельного мешка. Вырост передней кишки в это время полости еще не имеет, так как в самой передней кишке полость еще не образовалась. Поскольку между обонятельным мешком и выростом передней кишки расстояние очень невелико, то почти сразу же после своего образования хоанальный вырост обонятельного мешка оказывается соприкасающимся с хоанальным выростом передней кишки (стадия 38 - 39 у обыкновенного тритона, стадия 34 у испанского). На этой стадии развития хорошо видно, что вырост передней кишки даже немного больше хоанального выроста обонятельного мешка и таким образом составляет по меньшей мере половину будущего хоанального канала (рис. 26, А, Б). Строго говоря, соединение выростов происходит по наклонной плоскости, и поэтому материал хоанального выроста обонятельного мешка заходит более далеко на дорсальной стороне будущего хоанального канала, а материал хоанального выроста передней кишки на вентральной стороне этого канала.
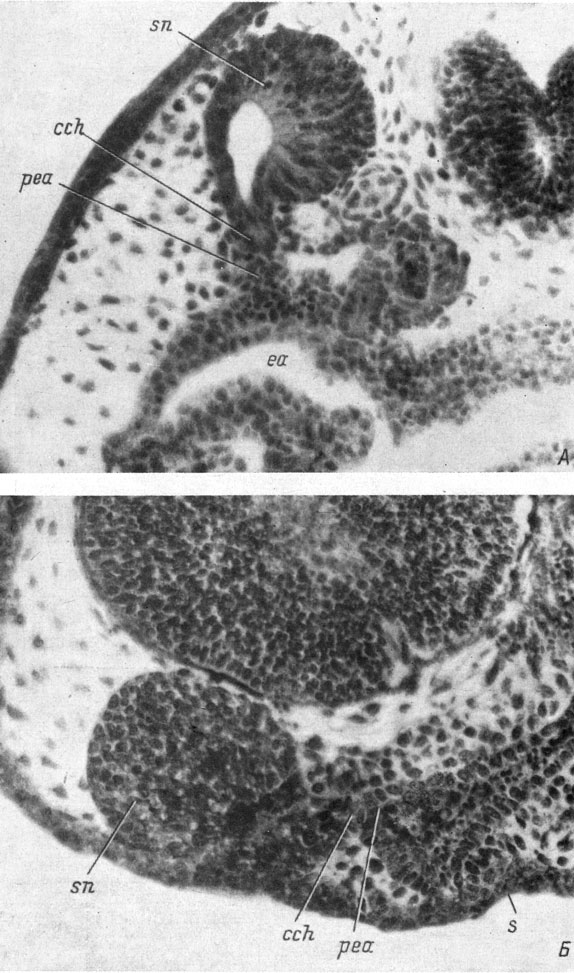
Рис. 26. Фронтальный срез головы личинки Pleurodeles waltlii длиной 10 мм, стадия 34 (А) и сагиттальный срез головы личинки Hynobius keyserlingii длиной 10.5 мм (Б). (Увел. 160)
Подойдя вплотную друг к другу, выросты срастаются (стадия 39 - 40 у обыкновенного тритона, стадия 34 - 35 у испанского тритона). Полость обонятельного мешка продвигается дальше в хоанальный вырост, а со стороны кишки появившаяся в ней полость входит в хоанальный вырост кишки, образуя таким путем хоану. Хоана, следовательно, у хвостатых амфибий, как и у бесхвостых, возникает ранее образования сквозного хоанального канала. Поскольку полость передней кишки развивается в заполняющей ее желточной энтодерме, и эта же энтодерма входит во внутреннюю часть хоанального выроста кишки, составляющего теперь каудальную часть будущего хоанального канала, то собственно хоана, т. е. соединительное отверстие между органом обоняния и ротовой полостью, образуется в энтодерме. Это хорошо видно на рис. 27, на котором даны фотографии сагиттального среза головы личинки Ranodon sibiricus при малом (рис. 27, А) и большом (рис. 27, Б) увеличениях. Хоанальный вырост обонятельного мешка и вырост передней кишки уже срослись. Полость обонятельного мешка и полость передней кишки довольно глубоко заходят в выросты, но еще не соединились, потому что пока только наружный эктодермальный слой выроста кишки сросся с наружным слоем эктодермы хоанального выроста обонятельного мешка. Желточная же энтодерма, которая на этой стадии еще хорошо отличима, так как в ней сохранились отдельные желточные гранулы, продолжается из выстилки полости кишки на внутреннюю поверхность стенки каудальной части хоанального канала, представляющей собой вырост передней кишки, и, дойдя до границы с выростом обонятельного мешка, давшим ростральную часть хоанального канала, образует перемычку, разделяющую эти две части хоанального канала. Внутренние стенки хоаны сформированы, следовательно, только энтодермальными клетками, хотя сам хоанальный канал в основном состоит из клеток эктодермы, причем как обонятельного мешка, так и обкладочных клеток выроста передней кишки. Этот момент развития органа обоняния очень трудно уловим, так как полости обоих хоанальных выростов очень быстро соединяются и образуется сквозной хоанальный канал (стадия 39 - 40 у обыкновенного тритона, стадия 34 - 35 у испанского тритона). Вскоре после возникновения сквозного хоанального канала полость передней кишки, формирующаяся путем расхождения энтодермальных клеток и, видимо, частичной их резорбции, распространяется далее рострально и прорывается наружу, образуя тем самым ротовое отверстие. Таким образом, на всем протяжении развития ротовой полости хвостатых амфибий у них так и не возникает настоящей ротовой ямки, так как впячивание эктодермального материала будущей ротовой полости заменено его врастанием.
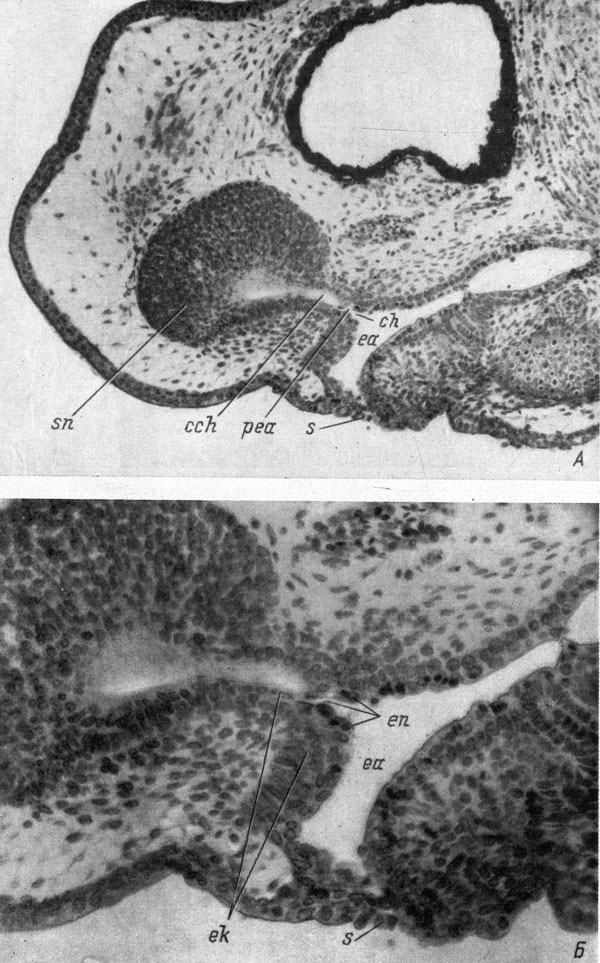
Рис. 27. Сагиттальный срез головы личинки Ranodon sibiricus длиной 18 - 19 мм (Л, увел. 80) и его деталь (Б, увел. 160)
Поскольку хоана и каудальная половина хоанального канала выстланы энтодермальными клетками, то образующийся на медиальной стенке хоаны клапан, очевидно, имеет энтодермальное происхождение. У Ranodon sibiricus клапан возникает сразу же вслед за образованием хоанального канала, у Hynobius keyserlingii он появляется несколько позже, у обыкновенного, кавказского подвида гребенчатого и испанского тритонов гораздо позднее, незадолго перед метаморфозом (закладка клапана у обыкновенного тритона появляется на стадиях 48 - 50 по Глюкзон, а формирование его заканчивается к стадии 57 - 58; у испанского тритона закладка и формирование клапана протекают быстрее и происходят несколько раньше - на стадии 44 - 45).
Только после образования хоанального канала, т. е. после соединения органа обоняния с ротовой полостью, начинается описанное М. Курепиной (1927) перемещение ноздрей с вентральной стороны морды на дорсальную и происходит сдвиг ротового отверстия в конечное положение (стадия 46 - 47 по Глюкзон у обыкновенного тритона, стадии 38 - 40 у испанского тритона). Оба этих несомненно связанных между собой процесса протекают очень быстро у высших хвостатых амфибий и значительно медленнее у Hynobiidae, особенно у Ranodon sibiricus. У последнего вида рот оказывается сдвинутым на конец морды, а ноздри соответственно на дорсальную ее сторону только у личинок длиной 29 - 33 мм, у которых уже имеется три пальца на задней лапе и начинает образовываться зачаток четвертого, что примерно соответствует стадии 56 - 57 по Глюкзон у обыкновенного тритона и стадии 51 у испанского тритона.
Таким образом, у всех хвостатых амфибий наблюдается одинаковый способ образования собственно хоаны и хоанального канала. В процессе развития орган обоняния и наружное носовое отверстие перемещаются из исходного положения на переднем конце морды по ее вентральной поверхности каудально к области ротового впячивания (рис. 77). Это перемещение имеет следствием очень близкое расположение обонятельного мешка и будущей ротовой полости по отношению друг к другу в момент образования хоанального канала и, следовательно, небольшую длину последнего. Хоанальный канал хвостатых амфибий обязан своим возникновением не только выросту обонятельного мешка, но и выросту передней кишки, растущих по направлению друг к другу. Факт образования каудальной половины хоанального канала из выроста передней кишки служит лишним доказательством в пользу отсутствия у хвостатых амфибий оро-назальной борозды и образующегося из нее эпителиального тяжа, так как в противном случае пришлось бы допустить, что оро-назальная борозда не достигает ротового впячивания, но тогда она и не может являться оро-назальной бороздой, поскольку не осуществляет первичного соединения между обонятельной и ротовой ямками. Вопрос мог бы стоять только о возможности образования ростральной половины хоанального канала из постназальной борозды, описанной Бертмаром (Bertmar, 1966a) у Hynobius retardatus, и он будет обсужден ниже (стр. 47).
Возникающий путем соединения двух выростов (обонятельного мешка и передней кишки) хоанальный канал состоит в основном из эктодермальных клеток, но внутренние стенки той его части, которая прилегает к передней кишке, выстланы энтодермальными клетками кишки, и поэтому собственно хоана с ее клапаном является энтодермальным образованием. О прорыве хоанального канала в кишку говорить не приходится, так как хоана образуется раньше чем наступает соединение полости обонятельного мешка с полостью передней кишки, которое происходит в срединной части хоанального канала, на границе срастания хоанальных выростов обонятельного мешка и передней кишки.
Начинающееся еще до образования хоанального канала перемещение ротового отверстия вперед продолжается и после этого момента и приводит в результате к конечному положению рта и обратному смещению наружных носовых отверстий не только в конечное, но и в дорсальное положение (рис. 77, Д, Е), причем у Hynobiidae этот процесс совершается гораздо медленнее, чем у более высоко организованных хвостатых амфибий. Такое более медленное и более явственное перемещение наружных носовых отверстий (как первоначальное - из конечного положения к месту ротового впячивания, так и последующее - от края рта на дорсальную сторону головы) именно у Hynobiidae - самых примитивных представителей современных хвостатых амфибий - является прямым указанием на вероятность подобного перемещения ноздрей у предков амфибий в процессе их филогенетического развития.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'