2. Развитие ноздрей и хоан хвостатых амфибий
У эмбрионов хвостатых амфибий обособление головной части от туловища начинается непосредственно вслед за замыканием нервной трубки. Первыми выпуклостями поверхностного рельефа головы являются выступающие глазные пузыри и выпуклость переднего мозга. Однако степень развития этих отделов головы неодинакова у различных видов.
Наиболее рельефно выступают выпуклости глаз и переднего мозга у обыкновенного тритона - Triton taeniatus (=Triturus vulgaris), испанского тритона - Pleurodeles waltlii - и сибирского углозуба - Hynobius keyserlingii (рис. 16, А - Б). Эмбрионы этих трех видов, особенно первого, довольно сильно пигментированы уже на ранних стадиях развития, и благодаря наличию такой пигментации легко определить местоположение обонятельной плакоды, хотя она еще не выдается над поверхностью покровов. Обонятельная плакода располагается в продольном углублении, создаваемом находящимися рядом выпуклостями глаза и переднего мозга. Поскольку выпуклость переднего мозга довольно резко отграничена от вентральной поверхности тела эмбриона, упомянутое углубление, или ложбинка, проходя между глазом и передним мозгом, спускается на вентральную сторону, к месту будущего ротового впячивания. Эту ложбинку легко можно принять за оро-назальную борозду, так как она проходит в том же самом направлении, в котором должна была бы идти последняя; однако никаких следов образования лицевых отростков обнаружить при этом невозможно, а наличие ложбинки совершенно очевидно обусловливается соседствующими выпуклостями глазных пузырей и переднего мозга. При дальнейшем развитии и увеличении расстояния между выпуклостями глаз и переднего мозга эта ложбинка, естественно, становится менее заметной и, наконец, исчезает (рис. 16, А′ - В′). Дольше всего она сохраняется у обыкновенного тритона (рис. 16, А′), у которого как раз наиболее резко выражены выпуклости как глаз, так и переднего мозга. У Pleurodeles waltlii она исчезает еще до появления оформленного ротового впячивания. Рисунки М. Курепиной (1927, рис. 1 и 2), на которых изображены эмбрионы Triton taeniatus и Hynobius keyserlingii с резко очерченными глубокими оро-назальными бороздами, совершенно не соответствуют действительности.
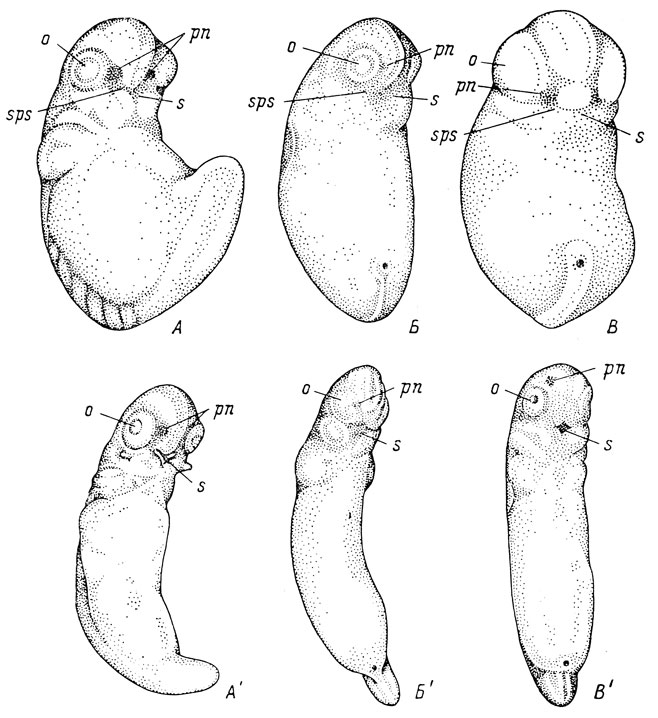
Рис. 16. Эмбрионы Triton taeniatus длиной 2 мм (А) и 3.3 мм (А'), Pleurodeles waltlii длиной 2.8 мм (Б) и 3.8 мм (Б'), Hynobius keyserlingii длиной 3 мм (В) и 5.3 мм (В'); вид с вентральной стороны и немного сбоку
У американского протея - Necturus maculatus (рис. 17, А) - и семи-реченского лягушкозуба - Ranodon sibiricus (рис. 18, Б) - выпуклости глаз очерчены довольно резко только с дорсальной стороны, а впереди полого переходят в слабо выдающуюся выпуклость переднего мозга. Тем самым между ними не имеется никакой ложбинки. Из-за более слабой пигментации по сравнению с описанными выше видами обонятельные плакоды при осмотре снаружи совершенно незаметны, и их положение между глазом и передним мозгом только угадывается. При дальнейшем развитии пологий переход между глазом и передним мозгом, с одной стороны, и между передним мозгом и вентральной поверхностью тела - с другой, сохраняется и никакой ложбинки, идущей от обонятельной ямки к месту ротового впячивания, так и не образуется. К моменту же появления ротового впячивания у Necturus maculates (рис. 17 А′) сильно развитые обонятельные доли переднего мозга создают между обонятельной ямкой и ротовым впячиванием выпуклость, а не углубление. У Ranodon sibiricus обозначившаяся обонятельная ямка также по-прежнему не имеет никакого соединения с наметившимся ротовым впячиванием (рис. 18, Б′).
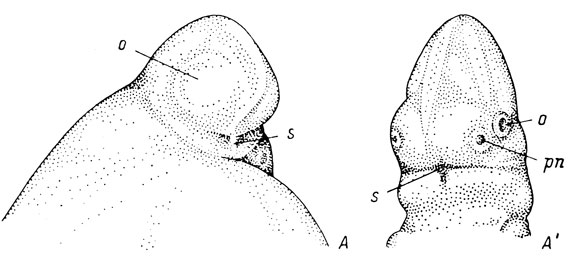
Рис. 17. Эмбрионы Necturus maculatus длиной 7.6 мм (А) и 10.5 мм (А') вид с вентральной стороны и немного сбоку
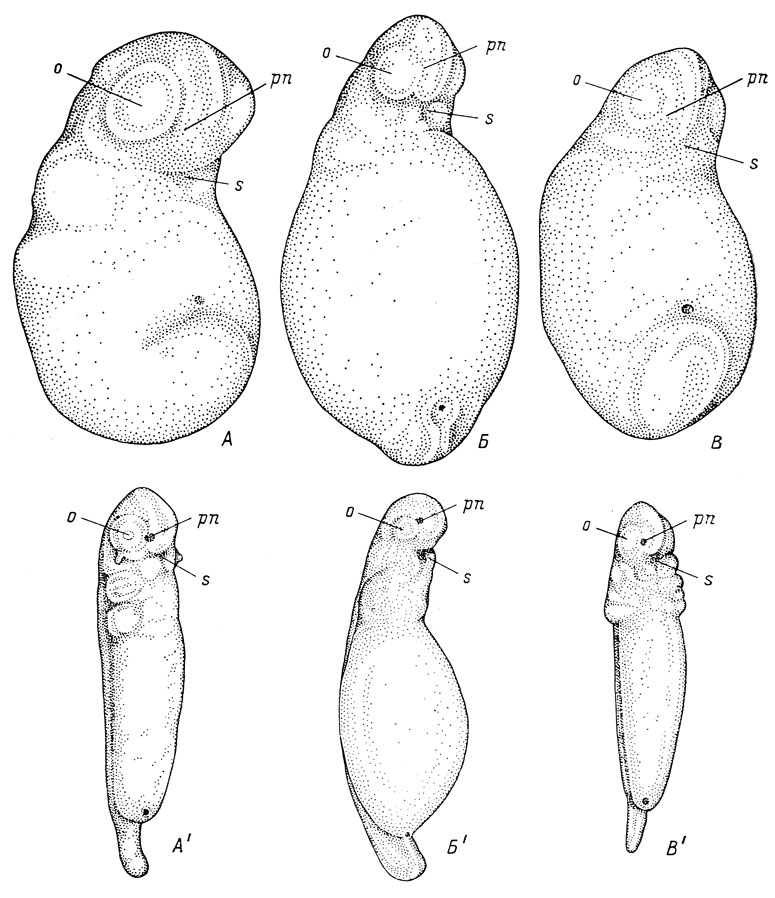
Рис. 18. Эмбрионы Triton cristatus karelini длиной 2.8 мм (А) и 5.6 мм (А'), Ranodon sibiricus длиной 5.9 мм (Б) и 8.5 мм (Б') и аксолотля длиной 2.5 мм (В) и 5.1 мм (В'); вид с вентральной стороны и немного сбоку
У кавказского подвида гребенчатого тритона - Triton cristatus karelini - выпуклости глаз и переднего мозга выражены чрезвычайно слабо (рис. 18, А). У аксолотля - Ambystoma sp. - выпуклости глазных пузырей почти совсем незаметны, а выпуклость переднего мозга выдается над поверхностью очень незначительно (рис. 18, В). Как следствие, у обоих видов нельзя обнаружить ничего похожего на борозду между обонятельной плакодой и местом будущего ротового впячивания. В дальнейшем, хотя выпуклости глаз и начинают очерчиваться более резко, они все же полого переходят в выпуклость переднего мозга и углубления между ними так и не образуется (рис. 18, А′, В′).
М. Курепина (1927) находит, что у аксолотля оро-назальные борозды выражены еще резче, чем у обыкновенного тритона и Hynobius keyserlingii. Изображенный ею эмбрион аксолотля обладает не только глубокими оро-назальными бороздами, но и сформированным ротовым впячиванием, хотя по стадии развития он моложе изображенного здесь на рис. 18, В′, у которого ротовое впячивание только начинает намечаться. Однако на всем протяжении развития эмбриона аксолотля от замыкания нервной трубки до образования ротового впячивания обнаружить у этого вида хоть какое-нибудь подобие продольного углубления, напоминающего оро-назальную борозду, как например у обыкновенного тритона или же H. keyserlingii, не удается. Напомню, что Баузенгардт (Bausenhardt, 1939), подтверждавший существование оро-назальных борозд у Alytes obstetricans, так и не смог найти их у аксолотля.
Таким образом, продольные углубления, расположенные между обонятельной плакодой и местом ротового впячивания, можно обнаружить только у тех видов хвостатых амфибий, эмбрионы которых одновременно обладают ярко выраженными выпуклостями глазных пузырей и переднего мозга, причем лишь на очень ранних стадиях развития, когда глаза и передний мозг расположены непосредственно рядом друг с другом. Отсюда становится совершенно ясно, что видимая в этих случаях продольная ложбинка, проходящая между обонятельной плакодой и местом ротового впячивания, не имеет ничего общего с настоящей оро-назальной бороздой других позвоночных, а ее появление на короткое время объясняется только понижением рельефа поверхности головы между двумя расположенными рядом выпуклостями глаза и переднего мозга. Кавагое (Kawagoe, 1932) был совершенно прав, считая эту ложбинку простым углублением эктодермы, не имеющим отношения к оро-назальной борозде и к образованию хоанального канала.
Кроме того, нельзя не принять во внимание следующее обстоятельство. Если допустить существование оро-назальной борозды у хвостатых амфибий, то она должна расцениваться как примитивный признак, общий у них с другими, обладающими хоанами, позвоночными. Исходя из сказанного, можно было бы ожидать, что эта борозда окажется наиболее ярко выраженной у примитивных видов, из них же в особенности у Ranodon sibiricus, икра которого, откладываемая подо льдом в горных ручьях, развивается чрезвычайно медленно по сравнению с икрой других видов хвостатых амфибий, а ведь именно фактор замедленного развития кладет М. Курепина (1927) в основу своего объяснения причин появления более резко выраженных оро-назальных борозд, описываемых ею у аксолотля. Однако у R. sibiricus вообще не появляется во время развития никаких признаков углубления, которое можно было бы принять за оро-назальную борозду, а ближайший к нему вид, также очень примитивный, Hynobius keyserlingii, показывает несколько иные отношения и по наличию продольного углубления между обонятельной ямкой и местом ротового впячивания располагается в одной группе с обыкновенным и испанским тритонами, находящимися значительно выше него в филогенетическом древе хвостатых амфибий. Очевидно, что образование указанной ложбинки является видовой вариацией развития, не зависящей от положения в системе. Все же, для того чтобы отрицать с полным правом роль этого углубления в образовании хоанального канала у хвостатых амфибий, необходимо проследить его дальнейшую судьбу.
По данным М. Курепиной (1927), оро-назальная борозда хвостатых амфибий исчезает с поверхности тела, погрузившись под эктодерму, и превращается в эпителиальный тяж, соединяющий обонятельную ямку с ротовым впячиванием. Этот тяж, изображенный ею на рисунке сагиттального среза головы H. keyserlingii, а также на реконструкции по сагиттальным срезам (1927, рис. 5 и 6), появляется, согласно ее описанию, после исчезновения с поверхности оро-назальной борозды. Исследуя не один, а серии сагиттальных срезов головы не только H. keyserlingii, но и других хвостатых амфибий на последовательных стадиях развития, вплоть до момента образования хоанального канала, легко убедиться в том, что между обонятельной плакодой и местом ротового впячивания начиная с самых ранних стадий развития этих образований действительно имеется утолщенный (за счет внутреннего сенсорного слоя) слой эктодермы, соединяющий их друг с другом и имеющий на отдельных сагиттальных срезах вид эпителиального тяжа. Однако просмотр всей серии срезов сразу же показывает, что в данном случае речь должна идти не об эпителиальном тяже, а о достаточно широком эпителиальном пласте, находящемся на ранних стадиях развития между органом обоняния и областью рта. Что же это за эпителиальный пласт и чему он обязан своим происхождением?
Выяснить этот вопрос можно с помощью графических реконструкций поверхности головы эмбрионов по поперечным срезам, проецируемым на сагиттальную плоскость. При этом оказывается, что эпителиальный пласт тесно связан с развитием системы боковых линий головы, или сейсмосенсорной системы. На рис. 19 изображены реконструкции поверхностных образований щечной области головы у двух личинок H. keyserlingii длиной 7 мм и 9.5 мм. Первая из этих личинок должна, по данным М. Курепиной, обладать эпителиальным тяжом, возникшим из оро-назальной борозды и соединяющим обонятельную ямку с ротовым впячиванием. Реконструкция показывает, что на всем протяжении покровов от обонятельного мешка до наружной границы ротового впячивания располагается толстый эпителиальный пласт, являющийся закладкой инфраорбитальной плакоды органов боковой линии (рис. 19, А). Ни для каких-либо других образований между обонятельной ямкой и ротовым впячиванием просто нет места. Поскольку на этих стадиях развития материал плакод органов боковой линии все время перемещается и их закладки очень лабильны, то они на протяжении всего развития связаны между собой, а на ранних стадиях закладки всех боковых линий передней половины головы вообще составляют единую предушную плакоду, лишь позже разделяющуюся на закладки отдельных линий (Медведева, 1961б). Закладка инфраорбитальной плакоды соединена также и с краями обонятельной ямки и ротового впячивания хотя и не такими массивными, как она сама, но все же достаточно утолщенными пластами эктодермы, которые на приведенных рисунках графических реконструкций обозначены точками (более тесное расположение точек показывает увеличение толщины пласта). Поэтому, несмотря на перемещение закладки инфраорбитальной плакоды, соотношения не меняются, и обонятельная ямка и ротовое впячивание остаются все время соединенными между собой утолщенным пластом эктодермы, который на отдельных срезах всегда будет иметь вид эпителиального тяжа. Даже после образования хоанального канала, когда он уже соединился с передней кишкой, можно увидеть на отдельных срезах этот "эпителиальный тяж", так как соединение утолщенным эпителиальным пластом органа обоняния и наружных краев ротового впячивания еще существует вследствие того, что развитие и перемещение инфраорбитальной плакоды еще не закончено (рис. 19, Б). Сказанное справедливо также и для более ранних стадий, на которых, по данным М. Курепиной, должна была бы существовать оро-назальная борозда, так как и в это время закладки всех плакод соединены между собой непрерывным утолщенным слоем эктодермы.
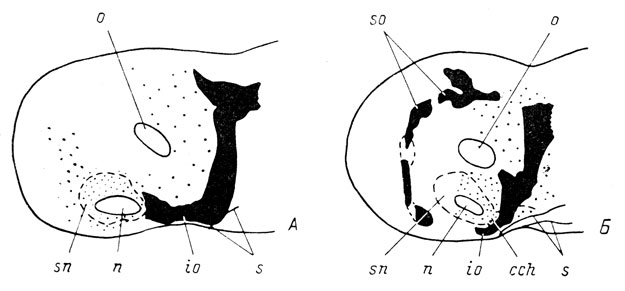
Рис. 19. Графические реконструкции поверхности головы эмбрионов Hynobius keyserlingii длиной 7 мм (А) и 9.5 мм (Б)
Видовые различия в образовании инфраорбитальной плакоды органов боковой линии сути дела не меняют. Реконструкция поверхности головы личинки Ranodon sibiricus длиной 10.8 мм (рис. 20, А) показывает, что хотя закладка инфраорбитальной плакоды не достигает обонятельной ямки, она продолжена утолщенным сенсорным слоем эктодермы, переходящим непосредственно в края ямки, благодаря чему осуществляется непрерывная связь органа обоняния с ротовым впячиванием посредством единого утолщенного эктодермального пласта. На более поздней стадии, у личинки длиной 15 мм (рис. 20, А), когда обонятельный мешок имеет уже небольшой хоанальный вырост и, следовательно, никаких следов поверхностного эпителиального тяжа не должно оставаться, соединение органа обоняния с ротовым впячиванием посредством утолщенного экто-дермального пласта продолжает сохраняться, и удачно проведенный сагиттальный срез по-прежнему покажет "эпителиальный тяж", который на этой стадии уже не должен был бы существовать.
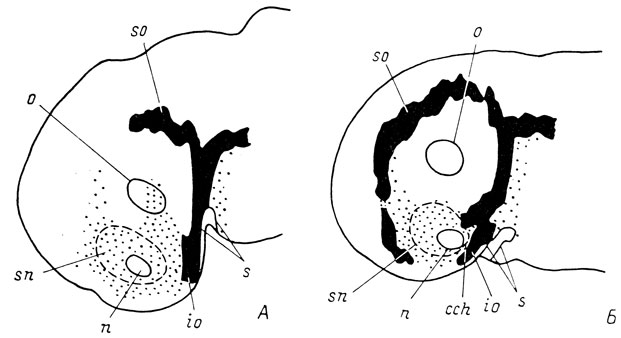
Рис. 20. Графические реконструкции поверхности головы эмбрионов Ranodonsibiricus длиной 10.8 мм (А) и 15 мм (Б)
Бертмар (Bertmar, 1966a) в своем недавнем исследовании образования хоан у Hynobius retardatus, H. nebulosus и Cryptobranchus (Megalobatrachus) japonicus также не нашел никаких оро-назальных борозд, но все же обнаружил небольшие углубления эктодермы, отходящие от обонятельных ямок по направлению к ротовому впячиванию, но не доходящие до него. Он назвал эти углубления постназальными бороздами и расценивает их как остатки исчезнувших оро-назальных борозд. Последнее заключение представляется сомнительным по причинам, изложенным в следующем разделе (стр. 47), но даже если Бертмар и прав, утверждая, что у некоторых хвостатых амфибий сохранились передние отрезки оро-назальных борозд в виде описанных им постназальных бороздок, то это обстоятельство все равно не имеет принципиального значения для окончательных выводов: постназальная борозда не осуществляет первичного соединения обонятельной ямки с ротовым впячиванием и, следовательно, не может образовать путем погружения в глубь тканей хоанального канала.
Таким образом, ни у одного из семи исследованных видов хвостатых амфибий, так же как и у бесхвостых, не наблюдается возникновения в процессе развития органа обоняния ни лицевых отростков, ни оро-назальной борозды как образования, дающего начало хоанальному каналу. На месте прохождения мнимой оро-назальной борозды у хвостатых амфибий располагается инфраорбитальная плакода органов боковой линии.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'