1. Развитие ноздрей и хоан бесхвостых амфибий
Поскольку суть разногласий в вопросе об образовании хоан амфибий заключается в том, существует ли у них оро-назальная борозда, как и у других наземных позвоночных, и принимает ли она участие в образовании хоанального канала и самих хоан, следует начать с описания внешней картины раннего развития органа обоняния. При макроскопическом осмотре головной части эмбрионов бесхвостых амфибий наиболее раннее появление рельефа обнаруживается у травяной лягушки - Rana temporaria. На стадии очень поздней нейрулы (нервные валики сомкнулись, но снаружи еще не срослись) при длине эмбриона 1.8 мм начинает намечаться присоска в виде идущих с боков двойных валиков, еще не соединяющихся посредине (рис. 10, А). Еле заметно обрисовываются глаза и жаберные возвышения. Поскольку не вполне сомкнувшиеся нервные валики приподняты над поверхностью головной части, они отграничиваются от последней чуть заметным углублением, передний участок которого (sps) может быть принят за оро-назальные борозды, но в действительности не является ими, так как еще нет ни обонятельных ямок, ни ротового впячивания. У эмбрионов той же длины, но чуть более поздних по развитию (нервные валики срослись полностью, однако шов еще виден) начинает намечаться ротовое впячивание и становятся чуть заметными обонятельные плакоды, но никакого углубления, соединяющего плакоду с ротовым впячиванием, нет, так как впечатление бороздок создавали приподнятые нервные валики, которые на этой стадии в результате полного срастания уже не возвышаются так над поверхностью головы, как это было на предыдущей стадии. На более поздних стадиях развития происходит образование обонятельных ямок и углубление ротового впячивания, но никаких следов соединяющей их борозды так и не появляется (рис. 10, А′). Что касается трех других исследованных видов (R. esculenta, Bufo viridis и Pelobates fuscus), то присоска, образующаяся первой изо всех рассматриваемых структур, возникает у них позже, лишь на стадии ранней хвостовой почки, причем в это время еще нет ни ротового впячивания, ни обонятельных ямок (рис. 10, Б - Г). Они появляются несколько позже, на стадии средней хвостовой почки, примерно одновременно с выпуклостями глаз и жаберными возвышениями, причем и в это время речь идет еще только об обонятельных плакодах. Никакого углубления, соединяющего их с ротовым впячиванием, ни на этой стадии, ни позже не появляется (рис. 10, Б′ - Г′).
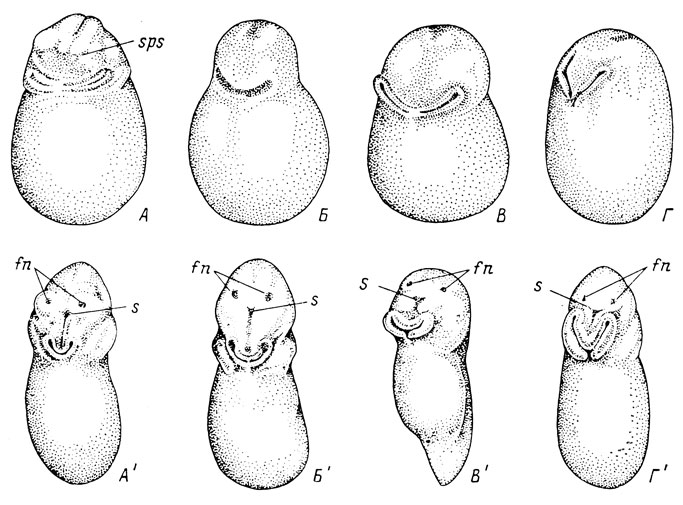
Рис. 10. Вид с ростро-вентральной стороны эмбрионов Rana temporaria длиной 1.8 мм (A) и 2.5 мм (A′), R. esculenta длиной 2 мм (Б) и 2.9 мм (Б′), Bufo viridis длиной 1.2 мм (B) и 2.4 мм (В′) и Pelobates fuscus длиной 2 мм (Г) и 2.7 мм (Г′)
У двух исследованных видов жерлянок (Bombina bombina s. Bombinator igneus и B. orientalis) также не возникает в процессе образования обонятельных ямок и ротового впячивания никаких соединяющих их борозд. У B. orientalis идут кпереди от ротового впячивания две расходящиеся под углом ложбинки, но они очерчивают выпуклость переднего мозга и проходят ростральнее обонятельных плакод. Во время образования обонятельных ямок выпуклость переднего мозга сильно сглаживается, и кроме самих ямок и ротового впячивания в передней части головы не наблюдается никаких углублений. У африканской шпорцевой лягушки - Xenopus laevis - на стадии ранней хвостовой почки от очень рано возникающего у этого вида ротового впячивания отходят вперед две ложбинки, отграничивающие выпуклость переднего мозга от выпуклостей глаз, и их-то и принял, по-видимому, Феске (Foske, 1934) за оро-назальные борозды, но в указанное время обонятельные плакоды снаружи еще не видны, а когда они появляются, то лежат сзади от описываемых ложбинок. В процессе дальнейшего развития обонятельные ямки отделены от ротового впячивания возвышением, а не углублением.
Поэтому вызывают недоумение рисунки М. Курепиной (1926, рис. 1, 2, 3), на которых изображены эмбрионы чесночницы (Pelobates fuscus) и зеленой лягушки (Rana esculenta) с широко расставленными нервными валиками, но имеющие вполне оформленную присоску и довольно глубокие обонятельные ямки и ротовое впячивание. Остается предположить, что использованный Курепиной материал по развитию зеленой лягушки не был пригоден для исследования такого рода вследствие того, что он был получен из засоленных болот, как об этом сообщает сам автор (Курепина, 1926, стр. 59), прибавляя, что "подобное изменение существования эмбрионов влияет замедляюще на ход их развития". По-видимому, в данном случае неблагоприятные условия существования вызвали не только замедление общего хода развития, но и задержку развития нервной системы, которая, как известно, первой реагирует на изменение условий. В частности, задержка замыкания нервной пластинки в трубку под действием солевых растворов была продемонстрирована еще опытами Гертвига над развивающейся икрой R. esculenta и R. fusca (Hertwig, 1895).
В результате подобной задержки срастания нервных валиков последние в соединении с уже развившимися присоской, ротовым впячиванием и обонятельными ямками могли дать картину ложных оро-назальных борозд, подобие которых наблюдается в норме у травяной лягушки, так как у этого вида перечисленные образования начинают развиваться несколько раньше, когда нервные валики еще не вполне срослись. Что касается зародышей зеленой жабы и чесночницы, развивавшихся в нормальных условиях, то здесь автор, по-видимому, был введен в заблуждение отдельными особями, уклонившимися от нормы и всегда встречающимися в кладке в небольшом количестве. Кроме того, у чесночницы на более поздних стадиях развития оро-назальные борозды могут быть симулированы тем, что присоска из-за своей большой величины у этого вида располагается очень близко к обонятельным ямкам и ротовому впячиванию, создавая тем самым небольшое углубление между ними (рис. 10, Г′).
Изучение серийных срезов развивающихся зародышей показывает, что раннее развитие обонятельного органа протекает очень сходно у всех исследованных видов, за исключением Xenopus laevis. Поэтому ниже дается описание развития органа обоняния у зеленой лягушки - Rana esculenta - с добавлением данных по другим видам и отдельно описание развития этого органа у Xenopus.
У эмбриона зеленой лягушки длиной 2.3 мм (стадия ранней хвостовой почки) обонятельный орган представлен парными обширными утолщениями, образованными в основном глубоким чувствующим (сенсорным) слоем эктодермы. Эти утолщения - обонятельные плакоды - соединены чувствующим слоем эпителия как между собой, так и с ротовым впячиванием. В области ротового впячивания чувствующий слой сильно утолщен, вдаваясь в глубь тела, и плотно прилегает к энтодерме передней кишки, составляя с ней единое целое. У эмбрионов длиной 3.5 мм исчезает постепенный переход чувствующего слоя обонятельной плакоды в чувствующий слой эпителия покровов, благодаря чему плакоды резко отграничиваются от покровного эпителия. Намечается впячивание центральной части плакоды и образование обонятельной ямки. У эмбрионов длиной 4.5 мм на задне-медиальном участке плакоды начинает обозначаться вырост, направленный каудально, совершенно сходно с описанным Гинсбергом у R. fusca (Hinsberg, 1901). Постепенно увеличиваются углубления обонятельных ямок и ротового впячивания. На всем протяжении развития сохраняется соединение обонятельных плакод с ротовым впячиванием посредством утолщенного чувствующего эпителия, действительно имеющего на срезе вид эпителиального тяжа, о котором писала М. Курепина (1926). Однако просмотр серии срезов сразу показывает, что это не тяж, а широкий пласт чувствующего слоя покровного эпителия.1
1 (Вопрос о природе этого пласта сенсорного эпителия будет подробно разобран на стр. 39 - 40 после изложения данных о наличии оро-назальной борозды у хвостатых амфибий.)
У эмбрионов длиной 5 мм задне-медиальный вырост обонятельной плакоды, превратившейся уже в обонятельную ямку, отверстие которой представляет собой первичную ноздрю, удлиняется каудально и несколько вентрально. Одновременно начинает обособляться круглый выступ в ее дорсальной части, направленный латерально, так называемый латеральный аппендикс. Ротовое впячивание углубилось настолько, что между ним и полостью передней кишки остается лишь тонкая перемычка, рото-глоточная мембрана, состоящая в своей центральной части только из одного ряда клеток. У эмбрионов длиной 5.5 мм обонятельная плакода превратилась в обонятельный мешок, и его задне-медиальный вырост, который теперь уже можно называть хоанальным, заканчивается на уровне дна ротового впячивания почти вплотную у его стенки, образующей в этом месте возвышение, как бы выступающее навстречу хоанальному выросту. У эмбрионов длиной 6 мм рото-глоточная мембрана еще имеется. Хоанальный вырост обонятельного мешка продолжается выше мембраны и подходит вплотную к кишечной трубке в месте, расположенном каудальнее первой, т. е. в области энтодермы. Срастание с кишкой еще не произошло, и клетки хоанального выроста хорошо отличаются от энтодермальных благодаря наличию в них пигмента. Этот момент соединения хоанального выроста обонятельного мешка с кишкой очень трудно уловим, так как именно в это время происходит прорыв и разрушение рото-глоточной мембраны. У эмбрионов длиной 6.5 мм уже нет никаких следов мембраны, и установить достоверно местоположение впадения хоанального выроста в кишечник невозможно. Поэтому для обнаружения описанных соотношений необходимы очень дробная фиксация и большое количество материала.
У травяной лягушки - R. temporaria - разрыв во времени между присоединением к передней кишке хоанального выроста и разрушением рото-глоточной мембраны несколько больше, чем у зеленой. Вырост подходит к энтодерме передней кишки у эмбрионов длиной 6 - 6.5 мм, у эмбрионов длиной 6.5 - 7 мм мембрана еще имеется. В месте присоединения хоанального выроста намечается небольшое углубление полости кишки внутрь выроста. Остатки рото-глоточной мембраны сохраняются еще у эмбрионов 7 - 7.5 мм длиной. Латеральный аппендикс также возникает относительно раньше, чем у зеленой лягушки. На стадии 6 - 6.5 мм он имеет вид оформленного шарообразного придатка, в котором уже есть полость.
У зеленой жабы - Bufo viridis - наблюдаются следующие соотношения. У эмбрионов длиной 3.7 мм хоанальный вырост подходит к передней кишке в области эктодермального ротового впячивания непосредственно рядом с рото-глоточной мембраной, т. е. в участке, пограничном между экто- и энтодермой, но не соединяется с ротовой полостью. На стадии 4.5 мм медиальный конец хоанального выроста уже сросся с энтодермальной частью передней кишки на довольно значительном расстоянии от еще имеющейся рото-глоточной мембраны. Полость передней кишки образует углубление внутрь еще плотного на остальном своем протяжении хоанального тяжа, клетки которого ориентированы в два слоя: тонкий - латеральный и толстый - медиальный. Прорыв мембраны происходит у эмбрионов длиной 5 мм; на этой стадии имеются как эмбрионы с сохранившейся мембраной, так и с ее остатками. Полость передней кишки довольно глубоко вдается в хоанальный вырост. Соотношение места впадения в переднюю кишку хоанального выроста обонятельного мешка и рото-глоточной мембраны показано на рис. 11. Латеральный аппендикс закладывается у эмбрионов длиной 3.7 мм, оформляется в виде шарообразного придатка при их длине 5 - 5.2 мм, полость в нем появляется у 6-миллиметровых эмбрионов.
У обоих исследованных видов жерлянок - Bombina bombina и B. orientalis - раннее развитие органа обоняния протекает весьма сходно.
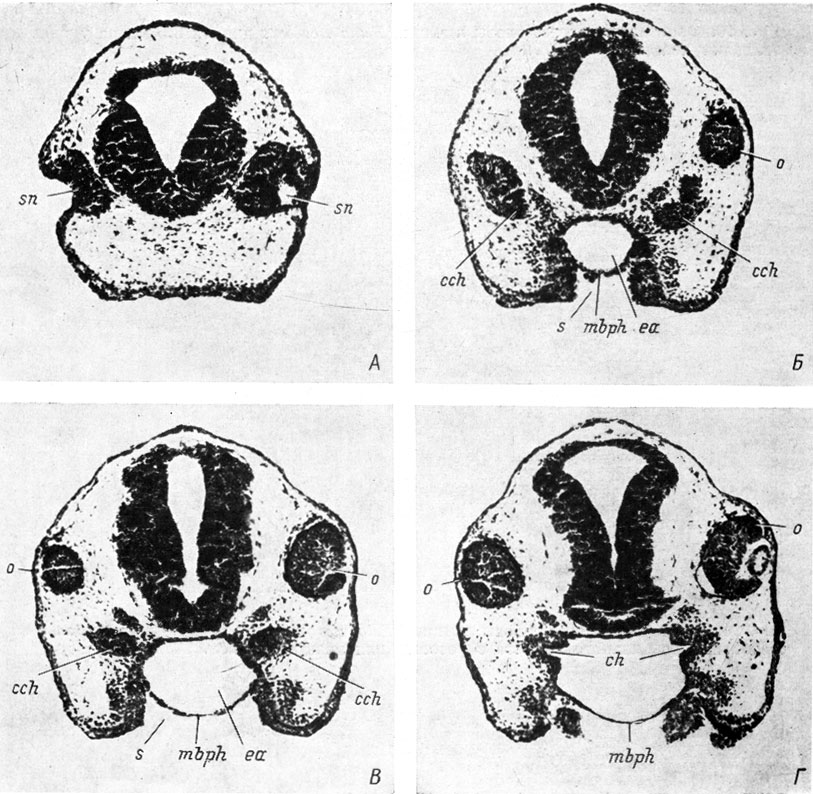
Рис. 11. Серия поперечных срезов головы личинки Bufo viridis длиной 5 мм (увел. 100). Срез на уровне: А - обонятельного мешка, Б - хоанального выроста, В - впадения хоанального выроста в переднюю кишку, Г - закладки хоаны
Уже на стадии поздней хвостовой почки в обонятельной плакоде, ясно отграниченной от эпителия покровов, появляется небольшая ямка, которая соединяется утолщенным подкожным слоем с ротовым впячиванием. Эктодермальный утолщенный слой, подстилающий ротовое впячивание, переходит без заметной границы в энтодермальную кишку. Энтодермальная часть этого общего рото-кишечного пласта заходит далеко вперед над ротовой ямкой (дорсальнее ее) и образует небольшой вырост по направлению к обонятельной ямке. У эмбриона B. bombina длиной 5.3 мм каудальный конец обонятельной плакоды, обладающей еще не очень глубокой ямкой, начинает удлиняться. У эмбрионов B. bombina длиной 5.8 - 6 мм имеется уже явственный вырост обонятельной ямки, направленный каудально, который у эмбрионов длиной 7 мм подходит к передней кишке над образовавшейся к этому моменту рото-глоточной мембраной, т. е. в энтодермальной части ротовой полости. У эмбрионов длиной 8 мм происходит соединение хоанального выроста обонятельного мешка с уже возникшим выростом энтодермальной части передней кишки совершенно так же, как у Ranidae и Bufo, причем в момент этого соединения вырост кишки уже имеет полость. Разрушение рото-глоточной мембраны у B. bombina осуществляется почти сразу же после образования соединения, у B. orientalis несколько задерживается.
В процессе дальнейшего развития происходит преобразование хоанального выроста обонятельного мешка в хоанальный канал. У эмбриона зеленой лягушки длиной 7 мм полость обонятельного мешка узкой щелью продолжается в хоанальный вырост, разделяя его на тонкую - латеральную и толстую - медиальную части. В это же время латеральный аппендикс приобретает полость. У эмбрионов длиной 8 мм полость хоанального выроста, являющаяся продолжением полости обонятельного мешка, удлиняется и довольно близко подходит к месту соприкосновения с выступом передней кишки. Полость последней заходит в этот выступ и ориентирована навстречу полости хоанального выроста. Направление сближающихся друг с другом полостей, представляющих собой узкие щели, не вполне совпадает. На рис. 12 можно видеть, что вместе с углублением полости передней кишки в конечный участок хоанального выроста входит энтодермальный материал. В вентральной части обонятельного мешка появилась закладка выступа, направленного медиально - якобсонова органа. У эмбрионов длиной 8.5 мм обе полости сливаются в единую полость хоанального канала. Благодаря тому, что при сближении выростов направление их полостей не вполне совпадало, углубление полости передней кишки впадает в полость канала ростральнее окончания последней. Вследствие этого участок ткани, разделявший конец полости канала и полость выроста передней кишки и состоявший как из эктодермальных, так и из энтодермальных клеток, обособляется в виде зачатка медиального клапана хоаны, который, таким образом, судя по его развитию, имеет смешанное экто-энтодермальное происхождение (рис. 13). У эмбрионов длиной 9 мм появляется второй, латеральный, клапан, расположенный каудальнее первого и, судя по его положению, возникающий только из энтодермальных клеток. При длине эмбрионов 9.2 мм вентрально-латеральная стенка обонятельного мешка, состоящая из индифферентного эпителия, начинает образовывать внутренний выступ, вдающийся в полость мешка - crista ventralis Ватанабе (Watanabe 1936a). Хоану закрывают вполне сформированные клапаны: более толстый - медиальный и более тонкий - латеральный. Вокруг ноздри образуется круговой валик, характерный для Ranidae. В якобсоновом органе возникает полость, являющаяся продолжением главной полости обонятельного мешка.
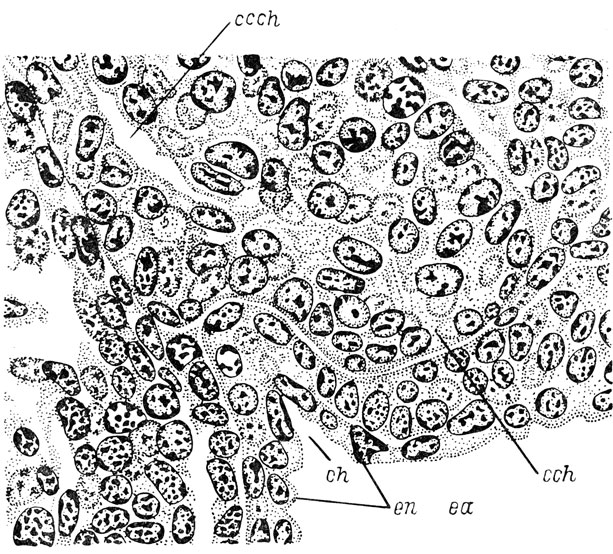
Рис. 12. Фронтальный срез головы личинки Rana esculenta длиной 8 мм на уровне соединения хоанального выроста с кишкой. (Увел. 740)
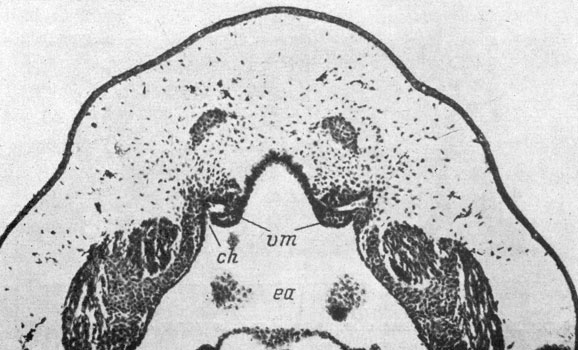
Рис. 13. Фронтальный срез головы личинки Rana esculenta длиной 8.5 мм на уровне закладки хоаны. (Увел. 70)
У травяной лягушки, зеленой жабы и обоих видов жерлянок образование полости хоанального канала происходит таким же способом, как описано выше. Закладка якобсонова органа возникает у эмбрионов травяной лягушки длиной 8 - 8.5 мм, хоана прорывается при длине их 9 - 9.5 мм, образование crista ventralis начинается при длине 10 мм. У эмбрионов длиной 10.5 - 11 мм полностью развиты оба клапана хоаны; при длине их 11 - 11.5 мм якобсонов орган приобретает полость.
У зеленой жабы якобсонов орган закладывается при длине эмбрионов 5 - 5.2 мм. В отличие от Bufo vulgaris, у которой, по данным Роведдера (Rowedder, 1937), имеется только медиальный клапан, у зеленой жабы образуются оба хоанальных клапана (5.2 мм), но в противоположность Ranidae медиальный вначале значительно больше латерального; у 6-миллиметровых эмбрионов они сравниваются по величине.
У жерлянок латеральный аппендикс запаздывает в развитии по сравнению с Ranidae и Bufo. Он возникает только у личинок длиной 8.5 мм (Bombina bombina) и 9 мм (B. orientalis), т. е. уже после срастания хоанального выроста с передней кишкой и резорбции рото-глоточной мембраны. В это же время образуется хоана путем продолжения полости кишки в хоанальный тяж, уже соединенный с кишкой; следовательно, она возникает в энтодерме. Клапаны хоаны развиваются не одновременно: сначала появляется медиальный (у B. bombina длиной 9 мм, у B. orientalis длиной 9.8 мм), а несколько позднее - латеральный. Якобсонов орган закладывается у B. bombina длиной 9 мм, а полость его начинает образовываться у личинок длиной 10 мм.
У чесночницы - Pelobates fuscus - формирование хоан и хоанального канала происходит несколько иначе, чем у других исследованных видов. На ранних стадиях развития (закладка обонятельной плакоды и преобразование ее в ямку) отличие состоит в том, что пласт клеток, соединяющий утолщенный чувствующий слой эпителия ротового впячивания с энтодермой передней кишки, имеет значительно большую протяженность в ростро-каудальном направлении, чем у остальных исследованных видов. На препаратах видно, что преобладающую часть этого пласта составляют энтодермальные клетки выроста передней кишки, отличающиеся от эктодермальных более крупными размерами и большей интенсивностью окраски. У эмбрионов длиной 5.2 мм начинается образование хоанального выроста обонятельной ямки и появляется закладка латерального аппендикса. У эмбрионов длиной 6 мм ротовое впячивание увеличивается на всю глубину экто-энтодермального пласта и очень близко подходит к полости передней кишки, в результате чего образуется рото-глоточная мембрана. На стадии 6.5 - 7 мм хоанальный вырост обонятельного мешка подходит вплотную к эктодермальному ротовому впячиванию, причем стенка последнего в месте соприкосновения образует небольшое возвышение. У эмбрионов длиной 7 мм в месте соприкоснования хоанального выроста со стенкой ротового впячивания последнее образует небольшое углубление в сторону выроста. У эмбрионов длиной 7 - 7.5 мм появляется закладка якобсонова органа. На стадии 7.5 мм хоанальный вырост обонятельного мешка срастается с возвышением стенки ротового впячивания впереди рото-глоточной мембраны, соединяясь, таким образом, не с энтодермальным, как у других видов, а с эктодермальным участком кишечного тракта; полость обонятельного мешка в это время немного заходит в вырост (рис. 14). У эмбрионов длиной 8 мм происходит прорыв рото-глоточной мембраны, увеличивается углубление полости ротового впячивания на проксимальном конце хоанального тяжа, на дистальном же его конце продвигается глубже в вырост полость обонятельного мешка. На стадии 8.5 мм полости соединяются и образуется сквозной хоанальный канал; начинают закладываться хоанальные клапаны. В это же время отмечается первое появление crista ventralis. У эмбрионов длиной 9 мм оба клапана развиты, причем латеральный по величине значительно больше медиального, как и у Ranidae. Размеры crista ventralis много больше размеров этого образования у других видов. Полость якобсонова органа возникает при длине эмбрионов 10.5 мм.
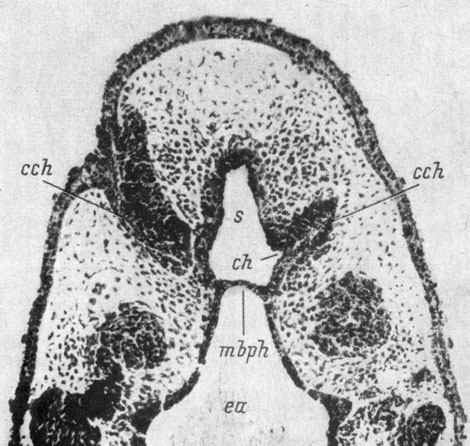
Рис. 14. Фронтальный срез головы личинки Pelobates fuscus длиной 7.5 мм на уровне соединения хоанального выроста с ротовым впячиванием. (Увел. 105)
Развитие хоан и хоанального канала шпорцевой лягушки - Xenopus laevis - стоит особняком и протекает весьма своеобразно по сравнению с описанными выше видами. На ранних стадиях развития (стадии 22 - 28 по Ньюкопу и Фаберу - Nieuwkoop a. Faber, 1956) массивные обонятельные плакоды заполняют все пространство между ротовым впячиванием, которое направлено прямо вперед, а не вентрально, как у других видов (начало образования впячивания - на стадии 26), передним мозгом и глазом и располагаются в вентральной половине самого переднего конца головы. Обонятельная плакода соединена утолщенным эктодермальным слоем с ротовым впячиванием, как это описал Феске (Foske, 1934). Рост плакод заметно отстает от усиленного роста головы, и их относительные размеры сильно уменьшаются (стадия 32). Начинает углубляться обонятельная ямка. На этой же стадии появляются рото-глоточная мембрана, так как углубление ротовой ямки идет очень интенсивно, а также небольшой округлый вырост на передне-медиальной стороне плакоды - якобсонов орган, который закладывается у X. laevis чрезвычайно рано, как это отметил впервые Феске. На стадии 33/34 мембрана очень утончается, а якобсонов орган приобретает полость.
На стадии 35/36 обонятельная ямка, по-прежнему лежащая в самой передней части головы и располагающаяся очень близко от эктодермального ротового впячивания, которое тоже занимает конечное, а не вентральное, как у других видов, положение, образует хоанальный вырост, находящийся чуть сзади и вентральнее якобсонова органа (рис. 15, А). Утолщенный эктодермальный слой, соединяющий обонятельную ямку с ротовым впячиванием, в это время еще сохраняется и никакого отношения к образованию хоанального выроста не имеет. Хоанальный вырост подходит к ротовой полости в области ее эктодермального впячивания, на этой стадии развития еще отделенного рото-глоточной мембраной от энтодермальной передней кишки. Ротовая ямка у X. laevis в отличие от всех описанных выше видов никакого выступа навстречу хоанальному выросту обонятельной ямки не образует. Эктодермальные клетки ротового впячивания несут в себе пигмент и поэтому хорошо отличимы от энтодермальных клеток передней кишки, не имеющих пигмента (рис. 15, Б). На стадии 37/38 описанные соотношения сохраняются, т. е. хоанальный вырост превратившейся уже в обонятельный мешок обонятельной ямки пока не сросся со стенкой эктодермальной части ротовой полости, а рото-глоточная мембрана еще имеется. На стадии 39 хоанальный вырост обонятельного мешка соединяется со стенкой ротовой полости, но мембрана, отделяющая эктодермальную часть ротовой полости от энтодермальной, уже резорбировалась. Однако несмотря на утрату этого важнейшего критерия, можно с уверенностью утверждать, что хоанальный вырост обонятельного мешка соединяется с ротовой полостью в ее эктодермальной части, поскольку в месте соединения все клетки ротовой выстилки пигментированы, а это присуще только эктодермальным клеткам (рис. 15, В). Феске (Foske, 1934), не имея столь подробного материала, какой был в моем распоряжении, все же предполагал эктодермальную природу хоан у Хеnopus на том основании, что обонятельные плакоды лежат далеко впереди, в области эктодермального ротового впячивания, и оказался прав в своем предположении. Однако он ошибся (опять-таки из-за слишком большого разрыва в исследовавшихся стадиях развития), согласившись с М. Курепиной, что хоанальный канал образуется из подкожного тяжа, соединяющего обонятельную и ротовую ямки, а не из хоанального выроста обонятельного мешка.
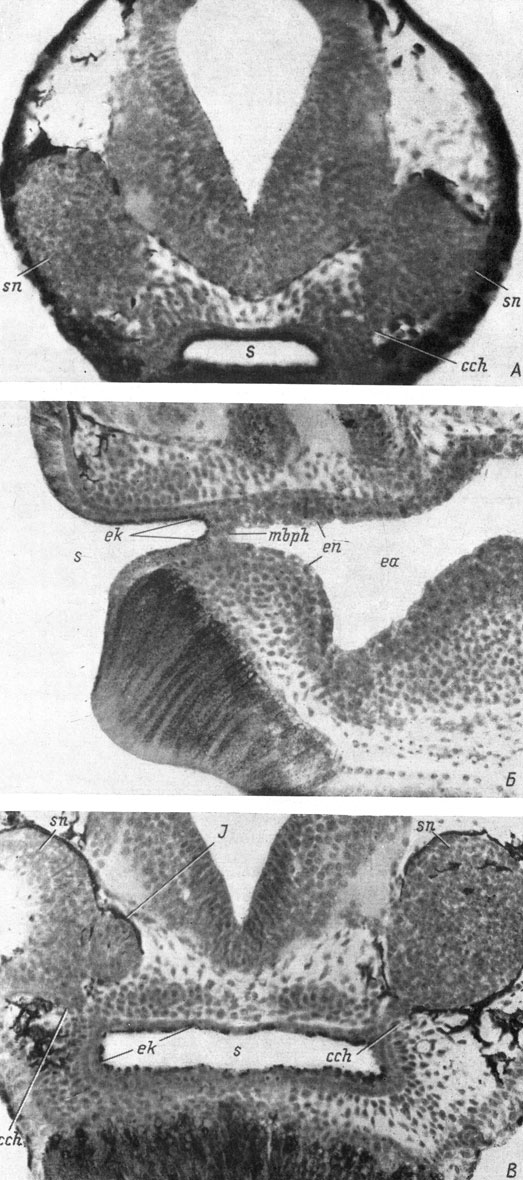
Рис. 15. Образование и соединение хоанального выроста обонятельного мешка с ротовым впячиванием у Xenopus laevis. А - поперечный срез головы личинки длиной 6 мм, стадия 35/36; Б - медианный сагиттальный срез головы личинки длиной 6.2 мм, стадия 37/38; Б - поперечный срез головы личинки длиной 6.5 мм, стадия 39 (увел. 190)
Дальнейшее развитие хоан и хоанального канала также протекает у Xenopus весьма своеобразно. Как уже упоминалось выше, в момент соединения хоанального выроста обонятельного мешка с ротовым впячиванием последнее не образует встречного выроста и соответственно ротовая полость не заходит в присоединившийся к ней конец хоанального тяжа, который в процессе дальнейшего развития еще долго остается плотным, так и не приобретая полости (со стадии 35/36 - личинка длиной 6 мм, до стадии 47 - личинка длиной 11.5 мм). Соответственно на всем протяжении этого периода развития не обнаруживается никаких признаков образования хоан. Лишь на стадии 47 у личинки длиной 13.5 мм начинают образовываться хоаны путем распространения полости рта в обращенный к ней конец все еще плотного на остальном своем протяжении хоанального тяжа. Сквозной хоанальный канал возникает только на стадии 48 (длина личинки 17 мм), а к стадии 49 (длина личинки 21 мм) медиальная стенка хоаны образует клапан. Этот медиальный клапан остается единственным, латеральный клапан хоаны у Xenopus не развивается.
На протяжении почти всего развития (возможно, причиной этого служит уплощенная форма головы, наблюдающаяся у личинок) вплоть до начала метаморфоза хоанальный канал и хоана располагаются непосредственно под ноздрей, а сам обонятельный мешок лежит сзади от ноздри и хоаны. Лишь в процессе метаморфоза, одновременно с преобразованием строения обонятельного мешка (развитие многочисленных добавочных мешков), хоана сдвигается назад и тем самым устанавливаются соотношения, характерные для других бесхвостых амфибий.
Таким образом, у всех исследованных видов бесхвостых амфибий способ возникновения хоанального канала одинаков: он образуется не из оро-назальной борозды, которой у бесхвостых амфибий не существует, а из выроста ткани самого обонятельного мешка. Рост хоанального выроста обонятельного мешка происходит в направлении эктодермального ротового впячивания. Расхождение в ходе развития наступает лишь в конечной стадии процесса: у чесночницы хоанальный вырост, подрастая к эктодермальному участку кишечника - ротовому впячиванию, соединяется с его стенкой, и хоаны образуются в эктодерме; у обоих видов лягушек, зеленой жабы и обоих видов жерлянок хоанальный вырост, дойдя до эктодермального ротового впячивания или до уровня рото-глоточной мембраны, продолжает расти до соединения с энтодермальным участком передней кишки и хоаны образуются в энтодерме. Ив том, и в другом случае образование хоаны начинается до прорыва полости хоанального канала в полость кишечной трубки за счет углубления полости передней кишки в прилежащий к ней конец хоанального тяжа.
Несколько иные отношения наблюдаются у шпорцевой лягушки. Хоанальный вырост обонятельного мешка подрастает к эктодермальному ротовому впячиванию и срастается с ним таким же образом, как у чесночницы, но образование собственно хоаны, т. е. соединяющего отверстия, задерживается и происходит много позже, когда распознать эктодермальную или энтодермальную природу клеток выстилки ротовой полости уже невозможно. Все же поскольку хоанальный вырост обонятельного мешка срастается с эктодермальным участком ротовой полости, тем самым он определяет первоначальное эктодермальное положение хоаны; более позднее образование собственно хоаны не меняет этого обстоятельства. У Xenopus не наблюдается и образования выроста ротовой полости для соединения с хоанальным выростом. С отсутствием выроста стенки кишечного тракта и связана, по-видимому, задержка в образовании хоаны. Этот факт также подтверждает, что хоана бесхвостых амфибий по своему возникновению относится не к хоанальному выросту обонятельного мешка, а к стенке кишечной трубки, причем она может быть как экто-, так и энтодермальной.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'