1. Строение и развитие органа обоняния рыб
При всех имеющихся разногласиях круг предков наземных позвоночных ограничивается костными рыбами (Osteichthyes). Из относящихся к костным рыбам двоякодышащих (Dipnoi), кистеперых (Crossopterygii) и лучеперых (Actinopterygii) рыб наиболее хорошо орган обоняния изучен у входящих в подкласс Actinopterygii хрящевых ганоидов (Chondrostei) и костистых рыб (Teleostei), а также у Dipnoi. У хрящевых ганоидов орган обоняния обладает более простым типом основного строения и больше приближается к исходному типу глубокой ямки, имеющемуся у низших рыб. У костистых рыб он отличается большим разнообразием размеров, формы и степени развития, причем наряду с усложненным строением органа встречаются и виды с почти полной его редукцией (О. И. Шмальгаузен, 1962). Это разнообразие безусловно объясняется экологическими различиями и обусловливается многочисленными частными адаптациями к внешней среде, которые соответственно не могут иметь глубокого филогенетического значения.
Орган обоняния костных рыб располагается на дорсальной стороне головы (исключение составляют Dipnoi в связи с особенностями развития органа), но положение это вторичное, так как у хрящевых рыб он находится на вентральной стороне, а у костных перемещается с вентральной стороны на дорсальную в процессе эмбрионального развития (Гавриленко, 1910; Reinke, 1936; О. И. Шмальгаузен, 1962; Teichmann, 1964, и др.). В своем наиболее простом виде орган обоняния лучеперых представлен более или менее округлым мешком, открывающимся на поверхности головы двумя отверстиями: передней (вводной) и задней (выводной) ноздрями (рис. 1). Как правило, вода входит в орган обоняния через переднюю ноздрю и выводится через заднюю, однако среди костистых рыб существуют виды, у которых обе ноздри функционируют попеременно то как вводные, то как выводные отверстия (Pipping, 1926, 1927). Слизистая органа обоняния образует складки, которые располагаются в форме розетки. Существуют различные типы розеток, наиболее распространено параллельное, радиальное, стреловидное и билатеральное положение складок (в последних двух случаях складки отходят от продольной септы). Вторичная складчатость, характерная для хрящевых рыб, у костистых отсутствует, за исключением щуки (О. И. Шмальгаузен, 1962). Обонятельные складки служат местом сосредоточения чувствующего обонятельного эпителия, остальная поверхность носовой полости выстлана индифферентным эпителием.
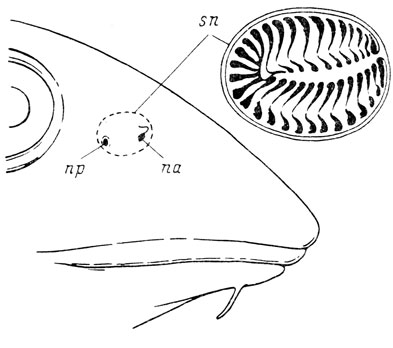
Рис. 1. Орган обоняния Gadus morrhua. По: Matthes, 1934
У многих костистых рыб, помимо главной носовой полости, имеются соединенные с нею один или несколько добавочных боковых мешков, не содержащих обонятельного эпителия. Предполагается, что функция их заключается в следующем: расширяясь и сужаясь при дыхательных движениях рыбы, они способствуют ритмической смене воды в носовой полости, а также, возможно, удалению из нее попавших частиц ила (Matthes, 1934). У некоторых видов драконообразных рыб добавочные мешки, по-видимому, могут приобрести связь с ротовой полостью посредством отверстия в нёбе (Atz, 1952a). Подобная связь самого обонятельного мешка с ротовой полостью у Astroscopus, описанная впервые Дальгреном (Dahlgren, 1908, 1927), рассматривается Атцем как вторичная, поскольку у родственного рода Uranoscopus на соответствующем месте имеется добавочный мешок. У некоторых угрей, относящихся к семействам Echelidae и Ophtichthyidae, различные виды демонстрируют разную степень приближения задних ноздрей к ротовой полости, вплоть до расположения их под верхней губой, т. е. собственно в полости рта (Atz, 1952b). В том и другом случае явление это представляет собой интересный пример конвергенции с образованием хоан наземных позвоночных и в дальнейшем будет разобрано подробнее.
У Polypterus delhesi, представителя стоящих особняком многоперых рыб, систематическое положение которых до сих пор нельзя считать окончательно установленным (еще не так давно их относили к кистеперым рыбам; Ярвик считает, что они ближе стоят к последним, чем к Actinopterygii (Jarvik, 1955); Леман тоже находит нужным выделить их в отдельную группу (Lehman, 1959)), обонятельный орган (рис. 2) интересен тем, что его полость не только разделена на вестибулярную, расположенную вентро-латерально, и чувствующую, дорсо-медиальную, части, но сама чувствующая часть имеет дополнительный дорсальный отдел, расцениваемый как добавочный мешок, но обладающий в отличие от таковых костистых рыб складками с обонятельным эпителием (H. Marcus, 1930; О. И. Шмальгаузен, 1962). В качестве образования, аналогичного добавочным мешкам костистых рыб и со сходной функцией, Маркус рассматривает вестибулярную часть органа, стенки которой снабжены гладкой мускулатурой. Передняя ноздря Polypterus вытянута в трубку, выступающую наружу и образующую щупальце, задняя, также представляющая собой трубку, проходит под кожей и открывается на поверхности небольшой щелью. Таково же устройство ноздрей у единственной современной кистеперой рыбы - Latimeria chalumnae (Millot et Anthony, 1954, 1958). Трубки передней и задней ноздрей открываются общим отверстием в носовую полость, в которой находится обонятельная розетка, состоящая из пяти обонятельных сосочков (Millot et Anthony, 1965). Пфейфер на основании изучения нескольких видов Polypterus и Calamoichthys calabaricus считает, что обонятельный орган Polypteridae вообще во всех отношениях больше сходен с органом обоняния латимерии, чем с таковым Actinopterygii и Dipnoi (Pfeiffer, 1968, 1969), но Тейсен находит различия в строении носа Calamoichthys и лучеперых недостаточными для выделения Polypteridae в отдельный подкласс (Theisen, 1970). Обонятельный мешок латимерии не расчленен (рис. 3) и, как у всех Coelacanthiformes, не имеет хоан, в отличие от ископаемых рипидистий, обладавших хоанами, а также, по мнению Ярвика, якобсоновым органом и слезно-носовым протоком (Jarvik, 1942) и представляющих собой, таким образом, единственную группу рыб, орган обоняния которых можно рассматривать как исходный для сложного органа обоняния наземных позвоночных.
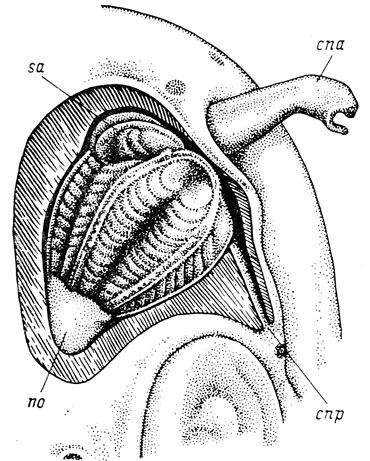
Рис. 2. Орган обоняния Polypterus delhesi. По: О. И. Шмальгаузен, 1962
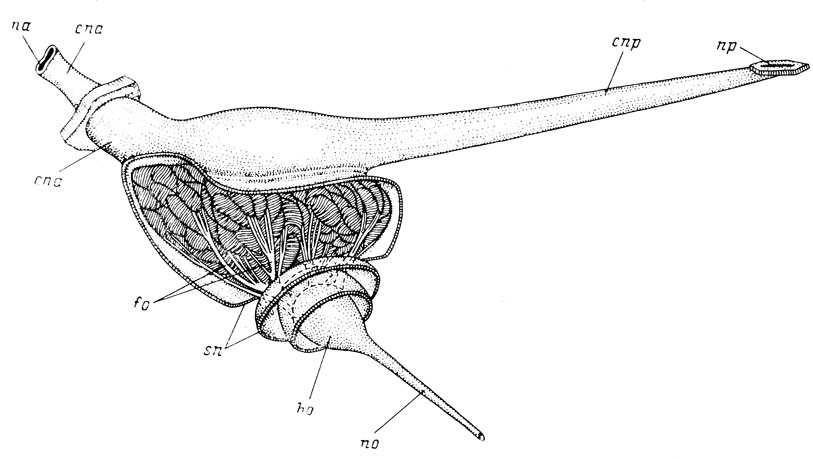
Рис. 3. Орган обоняния Latimeria chalumnae. По: Millot et Anthony, 1965
Как и во многих других отношениях, двоякодышащие стоят особняком от остальных костных рыб и в отношении органа обоняния. Прежде всего их обонятельный мешок обладает не только наружными ноздрями, но и внутренними, расценивавшимися ранее как хоаны. Но после исследования Neoceratodus forsteri Эллисом, оспорившим гомологию внутренних ноздрей Dipnoi с хоанами Tetrapoda на основании расположения первых во время своего возникновения снаружи от вторичной верхней губы и премаксиллярных зубов (Allis, 1932b), и в особенности после работы Ярвика (Jarvik, 1942), в которой он показал, что внутренние ноздри Dipnoi имеют совершенно иные отношения к частям хрящевой обонятельной капсулы и к прохождению r. maxillaris V, нежели хоаны истинных Choanata, сомнений в отсутствии у двоякодышащих настоящих хоан не остается. В последние годы эти данные подтверждены Бертмаром (Bertmar, 1965) и Томсоном (Thomson, 1965). Наружные, т. е. передние, ноздри взрослых Dipnoi хотя и не находятся в собственно ротовой полости, но все же расположены очень близко к краю рта, под вторичной верхней губой (рис. 4), и при закрытом рте снаружи не видны (Matthes, 1934)1. Соответственно положению ноздрей обонятельный мешок оказывается как бы перевернутым, и обонятельные складки располагаются не на дне, а на крыше носовой полости.
1 (Следует отметить, что у Protopterus передние ноздри хотя и находятся под верхней губой, но так как она довольно значительно выдается вперед нижней и приподнята над ней, то они открыты наружу и хорошо видны при взгляде спереди.)
Главная полость обонятельного мешка Dipnoi образует добавочные мешки, как медиальные, так и латеральные, из которых один, описанный впервые Керром у Protopterus (Kerr, 1910) и названный им латеральным дивертикулом, гомологизировался неоднократно с якобсоновым органом наземных позвоночных (Kerr, 1910; Broman, 1939b; Rudebeck, 1944, 1945; Schnakenbeck, 1960). Рудебек констатировал даже наличие у Protopterus рудиментарной добавочной обонятельной луковицы (к которой у наземных позвоночных подходят волокна якобсонова нерва), а также и самого рудиментарного n. vomero-nasalis, однако последующих подтверждений этого факта в литературе не имеется, и его достоверность вызывает большие сомнения (Parsons, 1967). В то же время Броман показал, что латеральный дивертикул у Protopterus и Neoceratodus имеет железистую структуру, а Фуллартон, указывая на сходство латерального дивертикула Protopterus и Lepidosiren с якобсоновым органом, сомневается в этой гомологии, поскольку упомянутый дивертикул образуется из дорсальной части стенки обонятельного мешка, а не из вентральной стенки, как у высших позвоночных (Fullarton, 1933). В недавно вышедшей подробной работе Бертмара (Bertmar, 1965) у представителей всех трех родов современных двоякодышащих рыб прослеживается преобразование мешкообразных дивертикулов, в том числе и гомологизируемого некоторыми авторами с якобсоновым органом, в обонятельные складки главной полости обонятельного мешка. Поэтому Бертмар считает, что никакой речи о наличии якобсонова органа у Dipnoi быть не может. Парсонс (Parsons, 1967, 1970а) указывает, что якобсонова органа нет ни у каких современных рыб. Как и у низших рыб, обонятельные складки носа двоякодышащих образуют дополнительно вторичные складочки (Matthes, 1934).
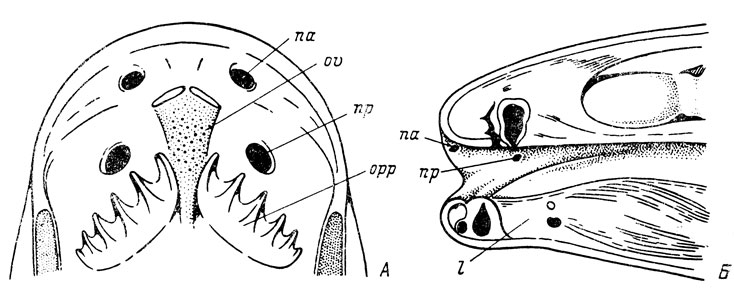
Рис. 4. Ноздри двоякодышащих рыб. А - Neoceratodus forsteri (по: Matthes, 1934); Б - Protopterus annectens (по: Паркеру из: Matthes, 1934)
Суммируя данные приведенного краткого обзора строения органа обоняния костных рыб, можно представить обобщенную схему строения этого органа. Он представляет собой замкнутый мешок, сообщающийся с внешней средой двумя отверстиями: вводным - передней ноздрей, и выводным - задней ноздрей. В некоторых случаях ноздри могут вытягиваться в довольно длинные каналы (трубки). Единственная группа рыб, у которых обонятельный орган имеет сообщение с ротовой полостью, образовав истинные хоаны, - это ископаемые кистеперые Rhipidistia. Вопрос о том, является ли отсутствие хоан у другой группы кистеперых, Coelacanthiformes, первичным или вторичным, пока не исследовано их развитие, остается открытым. У другой группы костных рыб, у которых также установилось сообщение органа обоняния с ротовой полостью, у Dipnoi, этот процесс протекал несколько иначе и привел к смещению обеих пар ноздрей в полость рта, причем передние ноздри все же имеют сообщение с внешней средой, а задние, несмотря на нахождение в собственно ротовой полости, топографически не соответствуют хоанам. Общая тенденция к смещению ноздрей в ротовую полость осуществлялась у двоякодышащих и у кистеперых различными путями, и здесь явно можно говорить о наличии у этих двух групп рыб параллельных процессов. Поэтому задние ноздри Dipnoi, в отличие от хоан Rhipidistia и наземных позвоночных, будут именоваться в дальнейшем изложении не хоанами, а внутренними ноздрями.
Упоминавшиеся выше внутренние ноздри некоторых угрей, исходя из того, что они наблюдаются лишь у отдельных представителей двух семейств костистых рыб, а у ближайших родственных групп (так же, как и у более примитивных Teleostei) ничего сходного обнаружено не было, явно представляют собой частную адаптацию и для рассматриваемой проблемы не могут иметь филогенетического значения. Хотя вопрос о филогенетическом значении внутренних ноздрей Astroscopus (сем. Uranoscopidae) столь же ясен, ему придется уделить несколько больше места в связи с недавней работой Бертмара (Bertmar, 1969). Утверждение этого автора, что внутренние ноздри Astroscopus, образовавшиеся путем соединения добавочного мешка с крышей ротовой полости, гомологичны хоанам рипидистий (а следовательно, и хоанам Tetrapoda), не имеет никаких научных обоснований. Единственным основанием для заключения такого рода могло бы служить сопоставление и согласование сравнительно-эмбриологических, сравнительно-анатомических и сравнительно-палеонтологических данных. Что касается первых, то ни развитие хоан рипидистий, ни развитие внутренних ноздрей Astroscopus нам не известно. Данные сравнительной анатомии показывают, что наличие третьей пары ноздрей, причем внутренних, ограничивается тремя видами рода Astroscopus, и даже у близких родов, не говоря уже о более отдаленных родственных группах, этих ноздрей нет (Atz, 1952a). Никаких палеонтологических данных, говорящих о наличии ряда предков Astroscopus, обладавших внутренними ноздрями, и приводящего к рипидистиям или их непосредственным предкам, также не имеется (сем. Uranoscopidae известно только с эоцена (Bertin et Arambourg, 1958)). Следовательно, как максимум, наличие внутренних ноздрей у рода Astroscopus может иметь филогенетическое значение только для выяснения возникновения этих образований внутри сем. Uranoscopidae. Этому вопросу, возможно, и не стоило бы уделять столько внимания, если бы не удивительный вывод Бертмара, сделанный им из своих построений: наличие хоан у Tetrapoda не имеет первостепенного значения в вопросе их происхождения, поскольку "таковые" обнаружены у Astroscopus. Естественно, что наличие внутренних ноздрей у одного рода из необъятного множества костистых рыб может рассматриваться лишь как интересный случай конвергенции в группе, стоящей очень далеко от эволюционного ствола Rhipidistia - Tetrapoda, не имеющий никакого отношения к происхождению наземных позвоночных и отнюдь не снимающий важнейшего филогенетического значения хоан.
Обонятельный мешок состоит из главной носовой полости, которая в некоторых случаях может расчленяться на вестибулярную и обонятельную части и иногда образует один или несколько добавочных мешков; однако пока не доказано ни у одного представителя костных рыб, что какой-либо из этих мешков гомологичен якобсонову органу наземных позвоночных. Наличие якобсонова органа, а также и слезно-носового протока у ископаемых кистеперых (Jarvik, 1942) тоже нельзя считать окончательно и безупречно установленным, так как оно предполагается только на основании существования соответствующих отверстий в обонятельной капсуле, поскольку мягкие ткани в ископаемом состоянии не сохраняются.
Слизистая обонятельного мешка рыб образует покрытые сенсорным эпителием обонятельные складки, собранные в розетки различных типов, что и является собственно чувствующей частью органа; выстилающий остальные участки органа обоняния индифферентный эпителий рецепторной функции не несет. К этому следует добавить, что сложных многоклеточных желез орган обоняния рыб не имеет, боуменовы железы также отсутствуют (Matthes, 1934). В слизистой находятся только одноклеточные бокаловидные железы (Matthes, 1934) и может продуцироваться секрет опорными клетками обонятельного эпителия (Fibak, 1953; Винников и Титова, 1957; Theisen, 1972).
Самая ранняя закладка органа обоняния позвоночных непосредственно связана с мозгом и представляет собой участок переднего медуллярного валика зародыша, находящегося на стадии нейрулы, как впервые было показано на рыбах А. Гавриленко (1910) и экспериментально доказано на амфибиях Цвиллингом (Z willing, 1940) и Купер (Cooper, 1943). Несколько позже О. И. Шмальгаузен (1950) обнаружила, что детерминация обонятельного зачатка у амфибий наступает еще раньше - на стадии гаструлы с большой желточной пробкой. Орган обоняния относится, таким образом, так же как и глаз, к первичным рецепторным органам, происходящим непосредственно из мозговой ткани (Винников, 1946), что делает понятным рост волокон обонятельного нерва от обонятельных рецепторов к мозгу. Поэтому обонятельные плакоды по своему происхождению и потенциям неравноценны другим сенсорным плакодам, не развивающимся непосредственно из нервной трубки.
У селахий обонятельные плакоды закладываются на дорсальной стороне головы, а затем смещаются на вентральную по боковой поверхности морды (Berliner, 1902; H. Marcus, 1930; Matthes, 1934). У Dipnoi обонятельные плакоды исходно дифференцируются на вентральной стороне головы, сдвигаясь в дальнейшем в ротовую полость (Greil, 1908; Matthes, 1934; Bertmar, 1965). Развитие обонятельного органа кистеперых неизвестно. У лучеперых рыб обонятельная плакода, закладываясь на вентральной стороне, позже смещается на дорсальную. Это перемещение происходит не через передний конец морды, как предполагали ранее (His, 1892; Keibel, 1893), а по боковой поверхности головы (Гавриленко, 1910; Reinke, 1936; О. И. Шмальгаузен, 1962). Последний автор полагает, что обонятельные органы оказываются на дорсальной стороне не вследствие изменения положения их зачатков, а в результате роста рострума впереди и вентральнее этих зачатков. Превращение обонятельной плакоды в обонятельную ямку, а затем в обонятельный мешок осуществляется путем роста и углубления плакоды в подлежащие ткани, ее инвагинации, а также либо расхождением и цитолизом чувствующего эпителия плакоды у одних групп, либо резорбцией поверхностного эпителия у других, либо обоими способами одновременно (О. И. Шмальгаузен, 1962).
Открывающееся наружу отверстие обонятельного мешка хрящевых рыб остается единым, но функционально оно разделяется на вводное и выводное отверстия благодаря образованию краями ямки лопастеобразных выростов, причем медиальная лопасть разрастается сильнее и перекрывает латеральную (рис. 5). В некоторых случаях она разрастается настолько, что достигает верхней губы и полностью закрывает заднее (выводное) отверстие - это наблюдается у многих акул и у большинства скатов (Matthes, 1934). При такой конфигурации медиальной лопасти образуется соединение между носовой и ротовой полостями в виде углубления, получившего наименование назо-буккальной (рото-носовой, оро-назальной) борозды (Allis, 1919).
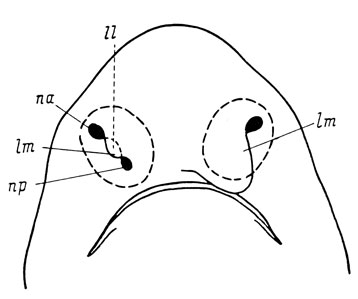
Рис. 5. Схема разрастания краев обонятельной ямки у селахий (слева - исходная форма, справа - тип Scyllium). По: Matthes, 1934
У костных рыб края обонятельной ямки, так же как и у низших рыб, образуют лопастеобразные выросты, растущие навстречу друг другу, но здесь эти выросты не перекрывают один другой, а срастаются, создавая таким образом носовую перемычку, или мостик, и разделяя тем самым единое отверстие обонятельной ямки на две, вполне изолированные друг от друга переднюю и заднюю ноздри. Сомнений в гомологии лопастеобразных выростов краев обонятельной ямки низших рыб с выростами, образующими носовую перемычку у высших, в настоящее время не возникает. Таким образом, обе ноздри костных рыб являются безусловными гомологами общего носового отверстия селахий.
У двоякодышащих рыб обонятельные ямки не сдвигаются на дорсальную поверхность головы, а остаются, как говорилось выше, на ее вентральной стороне. Первоначально они открываются наружу (Kerr, 1910; О. И. Шмальгаузен, 1962; Bertmar, 1965), но по мере образования ротовой полости и разрастания нижней челюсти вперед оказываются открывающимися в ротовую полость. Образование передней и задней ноздрей происходит точно так же, как у других костных рыб, путем срастания средних частей латерального и медиального краев первичного носового отверстия. В момент образования носового мостика передняя ноздря еще открывается наружу, тогда как задняя уже целиком находится в ротовой полости (Bertmar, 1965). В дальнейшем, по мере разрастания вторичной верхней губы, передняя ноздря прикрывается ею, но все же полностью в ротовой полости оказывается только задняя ноздря.
Последующее усложнение органа обоняния идет по пути образования складчатости чувствующей части обонятельной слизистой и формирования в ряде случаев добавочных мешков за счет выпячивания стенок главной полости. Многочисленные и разнообразные добавочные мешки костистых рыб не имеют чувствующего эпителия, образуясь в результате разрастания вестибулярной части органа, выстланной индифферентным эпителием (О. И. Шмальгаузен, 1962). Иначе обстоит дело у Dipnoi и, по-видимому, у Polypterus, развитие обонятельного органа которого не известно. У Polypterus имеется добавочный мешок, несущий складки обонятельного эпителия и располагающийся в ростро-дорсальной части обонятельного мешка (рис. 2), у двоякодышащих рыб прослежено развитие нескольких таких дивертикулов, в некоторых случаях подвергающихся последующей резорбции (Broman, 1939b). Однако, по недавним наблюдениям Бертмара (Bertmar, 1965), все появляющиеся в процессе развития органа обоняния Dipnoi дивертикулы в конечном итоге входят в состав главной полости обонятельного мешка и представляют собой только зачатки его отдельных обонятельных складок.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'