Пространственная структура популяции
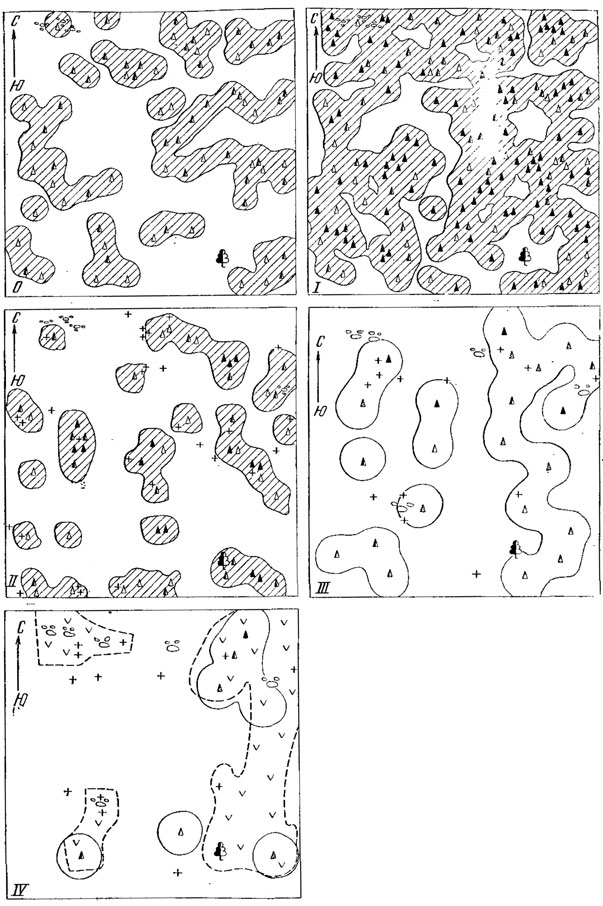
Рис. 88. Распределение особей прыткой ящерицы внутри одного поселения в разные сезоны в годы (данные и рисунок М. В. Тертышникова)
Точные данные о пространственной структуре значительного числа популяций отсутствуют, поскольку для прыткой ящерицы, как и для всех позвоночных, практически нет сведений о границах популяций (Тимофеев-Ресовский и др., 1973). Будучи достаточно тесно связанными с определенными пригодными для жизни местообитаниями и обладая сравнительно незначительным радиусом индивидуальной активности (порядка 5 - 15 м), группы особей прыткой ящерицы внутри популяции оказываются распределенными достаточно неравномерно. Эта неравномерность населения определяется неравноценностью отдельных участков как большой территории, так и внутри даже одного биотопа. Сплошной тип поселений свойствен лишь степным биотопам. Часто внутрипопуляционные объединения поселения ящериц оказываются "мозаичными", образуя как бы кружевной узор (рис. 88). Однако наиболее типичными оказываются поселения третьего типа - "ленточные", или "диффузные" (Тертышников, 1972б), как бы вкрапленные в относительно более обширные пространства, не имеющие постоянного населения. Примером таких поселений может служить распределение группировок прыткой ящерицы по разным биотопам Калужской области (рис. 89).
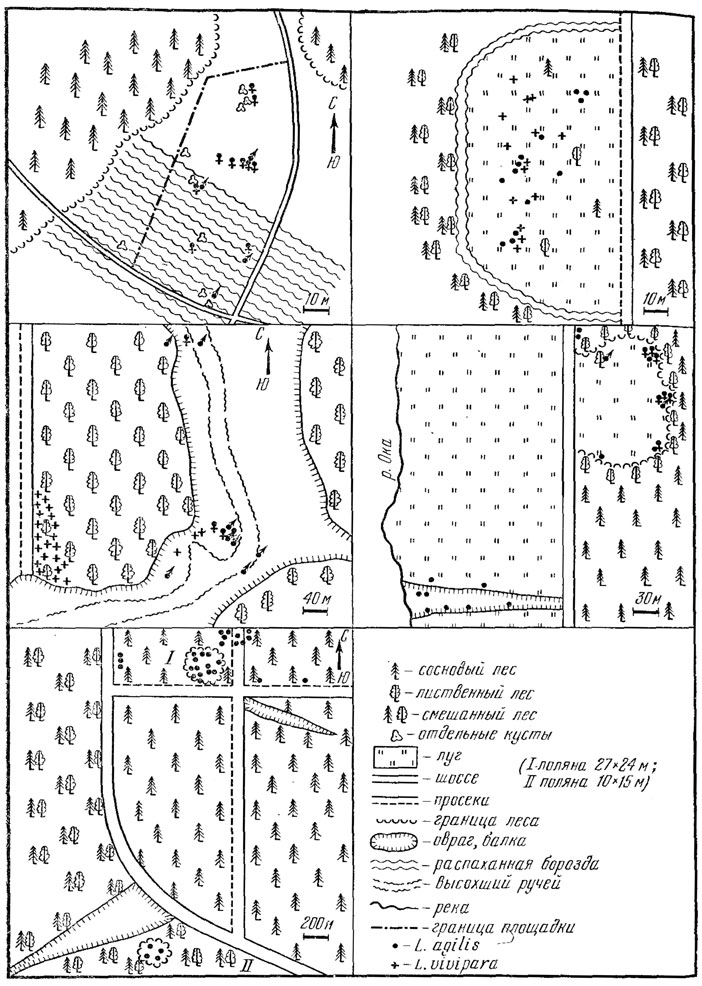
Рис. 89. Распределение групп особей прытких ящериц (семей, а может быть домов) в разных биотопах. Калужская обл., 1972 - 1974 гг. (данные А. Б. Стрельцова, рис. А. С. Баранова)
Следует учитывать, что границы между выделенными типами поселений оказываются подвижными: ежегодно, после выхода молодых, территория, занимаемая поселениями ящериц, резко увеличивается за счет распространения молодых особей по соседству с индивидуальной территорией самки (Тертышников, 1972). В годы высокой численности миграции ящериц, направленные обычно со стороны сплошных (т. е. более мощных) поселений в сторону мозаичных и диффузных поселений, также изменяют эти границы.
Внутри популяции взрослые ящерицы распределены неравномерно, обитая вблизи надежных укрытий (подробнее см. гл. V). Видимо, от наличия укрытий зависит и величина внутрипо- пуляционных группировок ящериц. Самые мелкие группировки состоят из нескольких (4 - 6 взрослых) особей, тогда как самые крупные из отмеченных группировок ящериц (временные скопления особей, например у воды, могут быть очень большими) могут достигать нескольких десятков особей (Щепотьев, 1948; Лукина, 1968б; и др.). Являются ли эти группировки особей семейными демами, надо решать в каждом конкретном случае специально, но такая вероятность не исключена. Во всяком случае существуют многочисленные наблюдения, указывающие как на длительность (до 2 - 3 лет) обитания определенных особей в конкретных норах и на длительные контакты между одними и теми же самками и самцами (Даревский, 1946; Лукина, 1966; Жаркова, 1973б; Тертышников, 1972; Щербань и др., сообщ.), так и на встречу самцов и самок на зимовке в одной норе (Яблоков и др., сообщ., район оз. Иссык-Куль), равно как и более частые встречи летом самцов и самок в одной норе. Интересным являются случаи встреч в одной норе нескольких взрослых ящериц (Стрельцов и др., сообщ.), большая часть которых оказалась взрослыми самками и только один - взрослым самцом (см. также рис. 88 - 89). Максимально тесные группы, состоящие из нескольких десятков особей, обнаружены в Западной Белоруссии (район г. Ковеля). Они обитали в единой колонии: отдельные норы были соединены между собой сетью сообщающихся ходов. Ориентировочная численность особей в такой колонии составляла не менее 25 - 30 взрослых животных (Баранов, Яблоков, сообщ.). Мелкие группировки типа демов обычно объединяются в более крупные в том случае, если подходящий для обитания ящериц биотоп занимает достаточно обширное пространство, например травянистый склон холма, длинный овраг, мощные заросли кустарников, лесополоса, просека в сосновом лесу и т. п. Численность таких вторичных группировок может достигать весьма значительных величин, порядка нескольких сотен взрослых особей, хотя чаще бывает меньше - несколько десятков особей. Так, например, на склоне холма площадью 300 м2 обитало несколько мелких групп общей численностью (за последовательные три года) 23 самца + 33 самки; 19 самцов + 31 самка; 16 самцов + 25 самок (Лукина, 1966б).
Итак, имеющиеся данные показывают, что внутри популяции ящерицы распределены неравномерно, небольшими (от нескольких до нескольких десятков взрослых особей) группами, приуроченными к благоприятным микробиотопам. Эксперименты показывают, что такие группы особей (вероятно, демы) могут существовать в относительно изолированном состоянии на протяжении жизни ряда поколений (Даревский, 1946; Боченко, сообщ.). Об этом же свидетельствует и значительная изолированность некоторых из таких демов: например, группа ящериц из 6 взрослых самцов и 6 самок, обитающая на склоне оврага на поляне в сосновом бору в Калужской обл., была удалена от ближайшего соседнего поселения ящериц на 2000 м, расстояние, многократно превышающее радиус индивидуальной активности в обычных условиях (см. рис. 89, Б). Лишь в периоды резкого подъема численности или миграций, связанных с неблагоприятными условиями, возможно преодоление этого пространственного барьера (Стрельцов, сообщ.).
Сказанное о внутрипопуляционной пространственной структуре относится в общей форме и к разным популяциям внутри ареала всего вида. Наблюдения в природе показывают, что у прыткой ящерицы существуют популяции, лишь незначительными изоляционными барьерами отделенные друг от друга и в совокупности образующие как бы сплошной тип поселения; такое тесное расположение популяций характерно для зоны оптимума, которая располагается в основном в степной зоне и частично в соседних (полупустынной и лесостепной) зонах. В других ландшафтах популяции ящериц оказываются распределенными мозаично, часто - ленточно (например, по поймам крупных рек или предгорным районам). Наконец, на севере и крайнем юге ареала популяции распределены диффузно. Так, например, южная граница ареала вида в районе 54 - 60° в. д. проходит по р. Эмбе (см. рис. 16). В этом районе в настоящее время происходит тесное соприкосновение полупустынного (левый берег) и сухостепного (правый берег) ландшафтов. Тщательное обследование показало, что единственная популяция L. agilis на протяжении нескольких десятков километров правобережья расположена неподалеку от пос. Жаркомыс, в зеленой долине размером около 1000х250 м, окруженной со всех сторон значительными песчаными дюнами. Но уже в 20 - 30 км севернее популяции ящериц образуют типичный кружевной узор, встречаясь недалеко друг от друга. Сходно и распределение ящериц Б горах Заилийского Алатау, в долине оз. Иссык-Куль. Отдельные группы ящериц (без специального стационарного исследования нельзя сказать, являются ли они малочисленными популяциями или представляют собой остатки каких-то миграционных волн, лишь временно обитающих на данной территории) встречаются по северному берегу озера, начиная от пос. Ананьево. Но первая настоящая популяция, с высокой численностью и постоянно обитающая на данном месте, встречается лишь в 35 - 57 км восточнее, по широкой балке и прилежащим окраинам полей у Тюбского залива. Здесь сразу же в непосредственной близости обитает несколько популяций (в том числе и интересные мелкие островные популяции), и эта большая группа популяций (видимо включающая в целом десятки популяций) совершенно изолирована от всего остального ареала прыткой ящерицы в Средней Азии: ни в восточной части Тюбской долины, ни в Кегенской долине ящериц нет. Ближайшие популяции обнаружены лишь на правобережье р. Или, за много десятков километров от оз. Иссык-Куль. В то же время прежние наблюдения показывают, что раньше ящерицы встречались и по левому берегу р. Или, т. е. можно предполагать, что изоляция группы популяций на Восточном Иссык-Куле не постоянная, а временная (Яблоков и др., сообщ.).
Приведенные в настоящей главе материалы показывают, что структура популяций прыткой ящерицы оказывается достаточно сложной (и в половом, и в возрастном, и в пространственном отношениях). Эта сложность многократно увеличивается в связи с подвижной "экологической" структурой популяции - изменениями активности отдельных возрастно-половых групп в разные сезоны года, и даже в разное время суток. Постоянно происходящие в природе микроэволюционные процессы, определяемые действием таких факторов, как естественный отбор, мутационный процесс, волны численности и развитие разных форм изоляции, постоянно нарушают казалось установившуюся структуру популяций, внося в нее элементы подвижности, текучести, динамики.
Структура популяции прыткой ящерицы оказывается несколько различной в разных участках ареала. Несомненно, существование географических различий в пространственной структуре популяций (преобладание сплошного типа поселений в оптимуме ареала вида и разного рода мозаичных и диффузных поселений на периферии ареала). Наконец, есть основания предполагать существование внутриареальной изменчивости половой и возрастной структуры популяции.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'