Активность
Методы учета активности
Наибольшее распространение в герпетологических исследованиях получил учет численности на ленточных пробах (трансектах). Эта методика достаточно подробно изложена в работах А. М. Андрушко (1936), Л. Г. Динесмана, М. Л. Калецкой (1952) и М. Ф. Тертышникова (1972б; 1973в) и заключается в следующем.
Прыткую ящерицу учитывают на ленте, ширина 'которой не превышает 2 м, а длина - от нескольких десятков до 1000 м. Каждая учетная лента должна находиться в пределах одного биотопа. В связи с неодинаковой плотностью поселения ящериц в разных частях биотопа 'рекомендуется многократная прокладка пробных лент в пределах изучаемого биотопа на расстоянии 5 - 10 м, причем маршрутный учет необходимо проводить при оптимальных условиях активности ящериц. В каждой географической зоне следует опытным путем определить эти условия. Так, для Ставропольской возвышенности установлено, что прыткая ящерица наиболее активна в период размножения - с середины мая до середины июня в первой половине дня при температуре воздуха 25 - 34°, почвы - 24 - 32°, относительной влажности воздуха 50 - 80% и скорости ветра от 0 до 12 м/сек.
Учет ящериц на ленточных пробах не позволяет определить их абсолютную численность, но остается незаменимым для получения ориентировочных данных о численности и биотопическом распределении этих животных и позволяет решать некоторые вопросы, связанные с их активностью и пр.
Для установления абсолютной численности следует закладывать изолированные площадки, размер которых должен быть равен 300 м2. Понятно, что изолированную площадку необходимо закладывать в пределах одного биотопа и отлавливать на ней ящериц в указанные выше сроки максимальной суточной и сезонной активности при наиболее благоприятных погодных условиях. Несмотря на трудности, связанные с оборудованием площадки (на ее закладку, включая изоляцию заборчиком из рубероида или полиэтиленовой пленки, требуется 6 - 8 человеко-часов), данный метод позволяет за первые три дня работы определить абсолютную численность ящериц.
Естественно, что для определения оптимальных погодных условий для ящерицы необходимо в процессе предварительной работы измерять влажность приземного слоя воздуха, температуру почвы, скорость ветра, температуру тела ящериц, фиксировать наличие осадков.
Суточная активность
Общий ход суточной активности
Известно, что прыткая ящерица имеет дневной цикл активности (Hoffmann, 1955, 1957; Тертышников, 1972б, и др.). По литературным данным, самый ранний выход ящерицы из убежища отмечен в 5 час. 45 мин. (Кутузова, 1971; Дагестан), а наиболее поздний уход - в 20 час. (Тертышников, 1972б; Ставропольская возвышенность). Летом это животное появляется из убежищ раньше, а уходит позже, чем осенью и весной. В зависимости от нарастания дневных температур у прыткой ящерицы может быть один - два пика активности, причем, весной и осенью их активность имеет один пик, а летом - два: утренний и вечерний с перерывом в самое жаркое время дня (Браунер, 1923; Красавцев, 1939; Либерман, Покровская, 1943; Даревский, 1946; Киселев, 1950; Хонякина, 1964; Щербак, 1966; Кутузова, 1971; Тертышников, 1972б; Боченко, Стрельцов, сообщ.). Основными факторами, влияющими на время выхода ящериц из убежищ, являются температура и осадки. Исследования М. Ф. Тертышникова (1972б) показали, что на Ставропольской возвышенности в хорошую погоду прыткая ящерица выходит из убежищ при интенсивности освещения около 200 лк. Из убежищ она появляется не сразу, а некоторое время лежит у входа, выставив голову наружу. В дальнейшем она полностью выползает из норы и греется в лучах солнца. При разной по температуре погоде прыткая ящерица затрачивает на обогрев от 0 до 160 мин. Достаточно прогревшись, ящерица начинает перемещаться в пределах индивидуальной территории. Интересно привести некоторые наблюдения М. Ф. Тертышникова за перемещениями взрослого самца прыткой ящерицы в разнотравно-злаковой степи на Ставропольской возвышенности. Выход животного из норы был отмечен в 7 час. 45 (мин. В течение дня он прошел 145 м, сделав 10 остановок разной продолжительности, на которые затратил всего 30 мин. Несколько раз самец возвращался в нору и провел в ней 6 час. Окончательный уход в нору был отмечен в 18 час. 30 мин. Таким образом, на поверхности это животное провело всего 4 час. 15 мин.
В плохую погоду выход ящерицы из убежища затягивается и может даже и не произойти. Перед заходом солнца ящерица тоже греется в лучах солнца у входа в нору. На ночь она прячется в убежище при интенсивности освещения от 21 000 до 280 лк. Ниже мы приводим рис. 82*, показывающий характер сезонной и суточной активности данной ящерицы в условиях Ставропольской возвышенности.
*(Форма обработки материала заимствована у Н. М. Окуловой (1969).)
Вес сеголетков принимали равным 1,5, молодых - 4,5, а взрослых - 12 г (см. гл. XV).
Для выяснения активности Lacerta agilia в лабораторных условиях был поставлен следующий эксперимент (Подмарев). Ящериц разделили на три группы. Первая группа обогревалась лампами накаливания круглые сутки. Вторая группа обогревалась с 8 час. утра до 18 час. вечера. Третья - контрольная группа - содержалась при комнатной температуре с естественным освещением и общим освещением лаборатории, по без дополнительного обогрева. Опыт проводится в течение года.
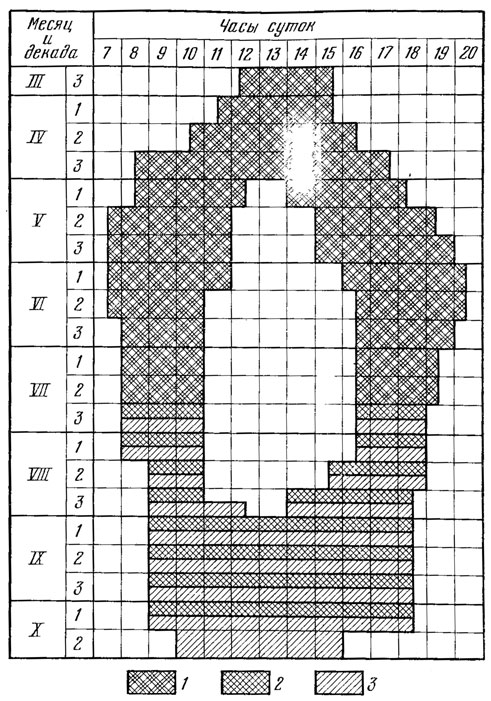
Рис. 82. Суточная и сезонная активность прыткой ящерицы на Ставропольской возвышенности. 1 - вся популяция, кроме сеголетков; 2 - единичные встречи взрослых и полувзрослых особей; 3 - сеголетки (по Тертышникову, 1972б)
При сравнении активности ящериц первой и второй групп различия в продолжительности суточной активности не были обнаружены. Наблюдения показали, что максимальная активность этих групп приходится на период с 11 до 14 час. дня. К 17 - 18 час. все ящерицы постепенно уходят в укрытия. С 8 - 9 час. утра ящерицы обеих групп выходят из укрытия и скапливаются у источника тепла, и к 11 час. все находятся в активном состоянии. В это же время замечены активное кормление и поиск пищи.
В контрольной группе, содержащейся без дополнительного обогрева, суточная активность выражена не так четко по времени, как у первых двух групп. Ящерицы контрольной группы редко выходят из укрытия на свет, плохо поедают корм в сравнении с первыми группами. Ящерицы контрольной группы подвержены различным заболеваниям (кожным, заболеваниям глаз и др.) и гибели. Следует отметить, что в первых двух группах этого эксперимента случаев заболевания ящериц не было.
За осенне-зимний период эксперимента существенных сезонных колебаний активности замечено не было. Лишь к весне активность выживших ящериц из контрольной группы стала приближаться к активности первых двух групп. В летний период все три группы по активности были одинаковыми.
Влияние погоды на суточную активность ящерицы. Температура тела и активность
По данным С. С. Либермана, Н. В. Покровской (1943; Московская область), температура тела активной прыткой ящерицы находится в пределах от 7 до 41° (рис. 83, см. рис. 57). В Крыму температура тела активной прыткой ящерицы варьирует от 9 до 40° (Щербак, 1962, 1966), а на Ставропольской возвышенности - от 14 до 36° (Тертышников, 1972б).
Наблюдения показывают, что прыткая ящерица активна при температуре воздуха от 4 - 7 (Стрельников, 1944) - 10,0 (Тертышников, 1972б) до 28,0 (Щербак, 1966) - 32,5° (Тертышников, 1972б). Изучая действие температуры среды1 на прыткую ящерицу в условиях Ставропольской возвышенности, М. Ф. Тертышников (19726) установил, что температура ее тела до 14° совпадает с температурой среды (рис. 84). При повышении температуры среды до 32 - 35° температура тела ящерицы оказывается выше на 0 - 7,5° или ниже на 0 - 14,5°. Однако при дальнейшем повышении температуры среды до 45° температура тела этого животного оказывается ниже ее на 0 - 7 или выше на 0 - 8°. Здесь же следует указать и на то, что молодые особи более пластичны к низкой температуре, но хуже переносят высокую температуру.
1(В данном случае и далее в тексте этой главы под "средой" понимаются почва и воздух.)
В различных регионах наибольшая активность ящериц проявляется при разной температуре. Так, например, в Московской области наибольшая активность ящериц зарегистрирована при температуре тела от 28 до 35° (Либерман, Покровская, 1943) (см. рис. 57, 83), а в Крыму - от 18 до 26° (Боченко, сообщ.). В Ставропольском крае это животное наиболее активно при температуре тела, равной 29,8, почвы - 29,0 и воздуха - 26,5 - 28,2° (рис. 84, А, Б). В этой связи интересно отметить, что максимальное количество кислорода прыткая ящерица в условиях данного региона поглощает при 30° (Тертышников, 1972б, 1973а).
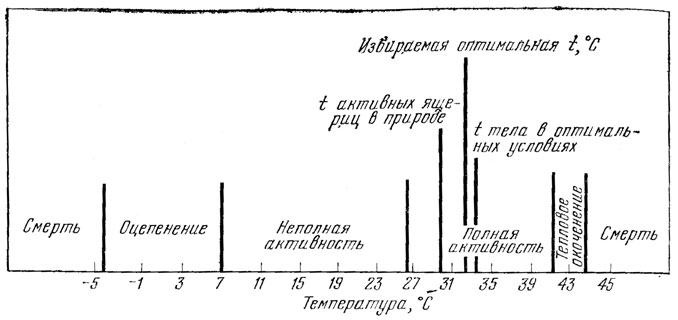
Рис. 83. Сводная схема температурных условий существования прыткой ящерицы в лесной и лесостепной зонах (по Либерман, Покровской' 1943)
При температуре среды и тела ниже 4 - 10° прыткая ящерица малоподвижна и в сезон активности обычно находится в убежище. По мере дальнейшего охлаждения почвы и воздуха ящерица теряет способность к активным перемещениям и впадает в оцепенение. С. С. Либерман и Н. В. Покровская (1943) установили, что в оцепеневшее состояние прыткая ящерица впадает при 7,5° (см. рис. 83). Использование микроклимата - одно из важнейших приспособлений ящерицы. В прохладные, но ясные дни она обогревается в лучах солнца и использует тепло нагретого грунта, прижимаясь к нему телом. Подобное поведение животного указывает на то, что солнечная радиация и тепловое излучение субстрата являются основными факторами, определяющими температуру его тела, и что температура воздуха имеет при этом меньшее значение. В подтверждение этого можно привести данные И. Д. Стрельникова (1944), из которых следует, что при температуре воздуха 4 - 7° температура тела прыткой ящерицы была равна 29,4 - 33,0°. Быстрое перемещение в холодную погоду также поднимает температуру тела ящерицы. И. Д. Стрельников (1934, 1944, 1959) отмечает, что после короткой пробежки температура тела прыткой ящерицы поднялась на 10°. Получив заряд тепла в условиях предварительного обогрева или после бега, ящерицы используют его, на некоторое время выходя из зависимости от низкой температуры окружающей среды. А. Д. Слоним указывает, что у рептилий не удается обнаружить адаптаций к холоду. Но, по всей видимости, именно наличие таких адаптаций обеспечивает активность прыткой ящерицы при низкой температуре среды и определяет возможность ее обитания высоко в горах на южной границе ареала.
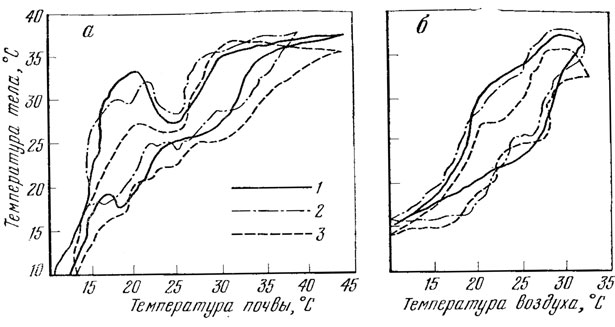
Рис. 84. Зависимость температуры тела прытких ящериц разных возрастных и половых групп от температуры почвы (А) и температуры воздуха (Б). 1 - взрослые самцы; 2 - взрослые самки; 3 - сеголетки (по Тертышникову, 1972б)
Выше было сказано, что, выходя из убежища, прыткая ящерица в прохладную погоду обогревается в лучах солнца. Обогревается она и после приема пищи. Когда ее тело сильно нагреется, ящерица ложится на брюхо, приподнимает конечности и хвост, покачивает головой и быстро моргает. В таком состоянии она остается на несколько минут, а затем быстро вскакивает и убегает в тень 'или же перебегает с места на место по освещенному солнцем участку. В таком же возбужденном состоянии ящерица бывает и при высокой температуре среды. Она быстро перемещается по субстрату, избегая прикасаться к нему телом, и вскоре во избежание перегрева уходит в тень от различных предметов, где температура среды ниже, чем на освещенных солнцем участках. В таких местах ящерица избегает также и прямого действия солнечных лучей, а температура ее тела снижается и определяется преимущественно температурой почвы. Кроме того, во время дневной жары прыткая ящерица способна влезать на ветки кустарников и стволы деревьев, где она избегает контакта с нагретым субстратом и где ветер снижает температуру ее тела. Весьма сходную реакцию прыткой ящерицы на действие высокой температуры описывают С. С. Либерман, Н. В. Покровская (1843) и В. В. Черномордников (1943, 1947).
Изложенный выше материал показывает, что при высокой температуре среды для прыткой ящерицы существуют определенные температурные пределы, ограничивающие пребывание этого животного на поверхности. По-видимому, для каждой географической зоны характерны определенные пределы. Для Ставропольской возвышенности, например, такими пределами, ограничивающими пребывание прыткой ящерицы на поверхности, являются температура почвы 32, воздуха - 30,6 и тела 36° (рис.85).
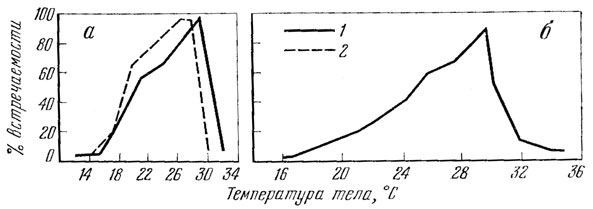
Рис. 85. Активность прыткой ящерицы в зависимости от температуры среды (а) и тела (б) по материалам наблюдения на стационаре в Ставропольском крае. 1 - температура почвы; 2 - температура воздуха (по Тертышникову, 1972б)
Температурные пределы, при которых способна жить прыткая ящерица, довольно значительны. Нижняя летальная температура равна - 4,9° (Либерман, Покровская, 1943). Верхняя же летальная граница варьирует в значительной мере в разных регионах.
В Московской области, например, она равна 44,7° (Либерман, Покровская, 1943) (см. рис. 83), на Ставропольской возвышенности - 41,0 - 41,6° (Тертышников, 1972б), а в горах Кавказа - 43 - 47° (Стрельников, 1944). Различия такого рода можно объяснить, по-видимому, наличием географической изменчивости терморегуляции ящерицы, которая, по мнению Л. Г. Динесмана (1949), имеет свои особенности в каждой географической зоне. В естественных условиях температура тела данной ящерицы не поднимается до указанных величин: у нее имеются элементы регулирования тепла, благодаря которым усиливаются или ослабляются действия того или иного фактора в каждой конкретной ситуации, а температура тела поддерживается на определенном уровне жизнедеятельности в течение дневной активности для обеспечения нормального протекания жизненных процессов. По И. А. Шилову (1962), эти элементы регулирования тепла не являются терморегуляцией: он называет их температурными адаптациями. Помимо перечисленных выше различного рода термотактических перемещений, к ним можно отнести еще испарение воды с поверхности тела и дыхательных путей.
Влажность, осадки и активность
О влиянии влажности среды на поведение прыткой ящерицы известно сравнительно немного. Так, по наблюдениям Н. Н. Щербака (1966), прыткая ящерица в Крыму активна в условиях относительной влажности воздуха от 56 до 100% при оптимальной ее величине, равной 56%. М. Ф. Тертышников (1972б) установил, что на Ставропольской возвышенности прыткая ящерица активна при 40 - 100%-ной относительной влажности воздуха, причем, наибольшая ее активность проявляется при 62 - 90%-ной влажности (рис. 86, А). По его же мнению, именно низкая влажность воздуха во второй половине лета вынуждает большую часть взрослых и полувзрослых ящериц впадать в летнюю спячку. В сопоставлении с температурой среды влажность имеет подчиненное значение, и ее высокий показатель не ограничивает выхода ящериц из убежищ.
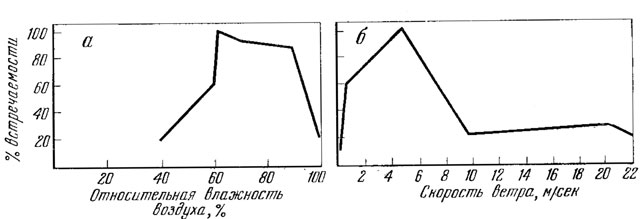
Рис. 86. Активность прыткой ящерицы в зависимости от влажности воздуха (а ) и скорости ветра (б). По Тертышникову (1972б)
Влажность среды в значительной мере формирует микроклимат и имеет большое значение в биотопическом размещении прыткой ящерицы: определяет наличие миграций (см. гл. V и IX) и выбор места для откладки яиц; установлено, что яйца могут нормально развиваться только при наличии 2,5 - 17,5% влаги в почве (Либерман, Покровская, 1943).
Непосредственное влияние на жизнедеятельность ящериц оказывают выпадающие осадки и стихийные бедствия, связанные с ними. Так, Л. Г. Динесман (1950) показал, что отсутствие ящерицы в нижней зоне дельты Волги вызвано ее неспособностью переносить паводки. По данным М. Ф. Тертышникова (1972б), периодические ливни в районе Прикалаусских высот на Ставропольской возвышенности весной и летом 1968 - 1969 гг. привели к гибели части популяций прыткой ящерицы, а отсутствие снегового покрова на некоторых участках не могло предохранить этих животных от гибели при действии низких зимних температур, когда в холодную зиму 1968 г. почва на таких участках промерзла на глубине до 100 см.
Сильный ветер снижает температуру тела ящериц и подавляет их активность при низкой температуре среды, но позволяет им быть активными при высокой температуре среды. По данным М. Ф. Тертышникова (1972б), прыткая ящерица на Ставропольской возвышенности активна при скорости ветра от 0 до 22 м/сек. Максимум активности ее отмечен при ветре, скорость которого равна 5 м/сек (см. рис. 86, Б).
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'