Наступление половой зрелости
Наступление половой зрелости можно объективно определить по развитию гонад, состоянию наружных половых органов и по поведению животного (спариванию). Эти методы пригодны только в период размножения. Поэтому более общими, хотя, несомненно, в известной степени основанными на косвенных данных методами являются определение половозрелости по возрасту и размерным признакам (весу, длине) тела и появлению вторичных половых признаков (развитие полового диморфизма по окраске).
У самцов прыткой ящерицы сразу же после выхода из спячки отмечается весенняя активация семенников, что приводит к сильному увеличению последних. Своего максимального размера они достигают в апреле-мае в зависимости от географического района исследования. В этот период семенники принимают цвет яичного желтка (Щепотьев, 1950). Изменение гонад в процессе размножения тесно связано с изменением жировых тел (Лукина, 1966б; Тертышников, 1972а, б; Симонян, 1973, и др.).
Жировые тела располагаются у нижнего края брюшной полости ящериц и имеют вид двулопастного образования беловато-желтого или ярко-желтого цвета. Известно, что у некоторых рептилий они состоят на 80 - 90% из различных липоидов (Dessauer, 1955) и в основном из арахидоновой и линолевой кислот (Чеченкин, 1954). В период максимального развития жировые тела имеют большие размеры, а при полном истощении они едва заметны.
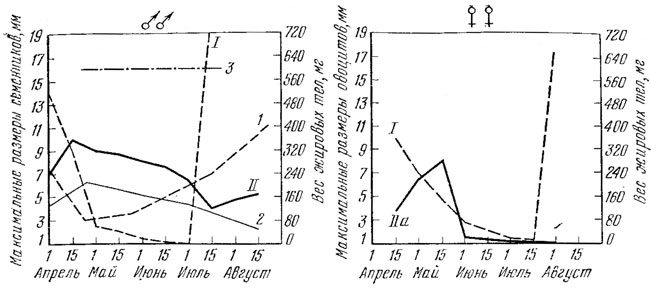
Рис. 66. Сезонные изменения веса жировых тел (I.1), максимальной длины семенников (II.2, нахождение зрелых сперматозоидов (3)и максимальные размеры овоцитов (IIа) у прыткнх ящериц Западного Предкавказья. I, II, а по Г. П. Лукиной (1966б); 270 самцов и 124 самки; 2, 3 - по М. Ф. Тертышникову (1972б)
Сезонная динамика жировых тел и гонад характеризует зависимость между питанием и размножением ящериц, что хорошо было прослежено Даревским (1960) в Армении. Для прытких ящериц Западного Предкавказья Г. П. Лукиной (1966) установлена взаимная зависимость изменения жировых тел и максимальных размеров гонад (рис. 66). При рассмотрении кривой видно, что у самцов после выхода из спячки длина семенников достигает значительной величины (до 10 мм), в то время как жировые тела расходуются целиком. С середины апреля до конца июня размер гонад постепенно уменьшается (до 4 мм), одновременно происходит быстрое нарастание веса жировых тел. За первую половину июля вес жировых тел увеличивается с 0 до 720 мг. К концу лета семенники снова несколько увеличиваются, что объясняется подготовкой их новому половому циклу. Применительно к данному виду ящериц летнее увеличение семенников было отмечено еще Н. Н. Щербаком (1962). В своей работе Н. Н. Щербак предполагает, что этот процесс связан, возможно, с понижением температуры и сокращением светового дня.
Аналогичные данные о динамике развития гонад самцов прыткой ящерицы приводятся и в других работах (Щепотьев, 1948; Краснова, 1961; Щербак, 1966 и др.).
Интересный факт приводит М. Ф. Тертышников (1972б) для прытких ящериц Ставрополья. Несмотря на то, что происходит прекращение сперматогенеза в летний период, зрелые спермин сохраняются на протяжении мая, июня и начала июля (см. рис. 66). По мнению И. С. Даревского (1967), подобная длительная половая активность самцов приводит к тому, что почти все половозрелые самки в популяции оказываются оплодотворенными; при соответствующих условиях в южных районах ареала возможна вторая кладка. При этом, по-видимому, не меньшее значение имеет установленный Д. И. Гениным (1955) факт длительного сохранения зрелых спермиев в половых путях самок.
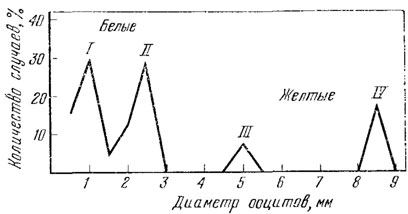
Рис. 67. Размеры ооцитов прытких ящериц в мае. Западное Предкавказье, 162 экз. (Лукина, 1966б)
Весной у пробудившихся самок в яичниках уже имеются несколько желтых и крупных (3 - 4 мм), а также матовых и мелких (1 - 2 мм) овоцитов.
Процесс развития яиц в яичниках половозрелых самок начинается с конца апреля. При этом у каждой самки развиваются 15 - 20 овоцитов, которые постепенно приобретают желтую окраску. Остальные овоциты отстают в росте и сохраняют белую окраску. Перед переходом в яйцеводы овоциты в яичнике представлены хорошо отличимыми по размерам четырьмя генерациями (рис. 67). Количество овоцитов в третьей генерации всегда мало (не более 4), а в четвертой оно, как правило, равно возможному количеству откладываемых яиц для данного вида.
Таким образом, четвертая генерация наиболее крупных овоцитов (8,5 мм) или ее какая-то часть развиваются дальше в яйцеводах. Овоциты третьей генерации редуцируются и в следующем сезоне, по-видимому в яйцеводах развивается вторая из отмеченных генераций.
Достигнув 8 мм в диаметре, яйца выпадают в полость тела, откуда переходят в яйцеводы (рис. 68). В это время и происходит оплодотворение. Развитие мелких овоцитов задерживается. В мае-июне готовые для откладки яйца, вес которых достигает 0,6 г, встречаются в яйцеводах половозрелых самок. У самок, окончивших размножение, перед уходом на зимовку в каждом яичнике уже имеется группа желтоватых овоцитов размерами до 3 мм.

Рис. 68. Расположение зрелых яиц в яйцеводах прыткой ящерицы. Фото В. И. Подмарева
У самок, так же как и у самцов, накопление и расходование жировых тел находится в тесной связи с динамикой развития половых продуктов (см. рис. 66).
Весной у самок при быстром созревании половых продуктов происходит истощение жировых тел. Однако их энергетических запасов видимо хватает на развитие яиц. По окончании размножения самки усиленно питаются и вес жировых тел быстро увеличивается, достигая максимума в конце июля. Часть вновь накапливаемых энергетических запасов расходуется на подготовку к очередному половому циклу.
Как видно (см. рис. 66), пробудившиеся от зимней спячки самки (апрель) содержат в яичниках 10 - 12 крупных ооцитов диаметром 3 - 4 мм и большое количество мелких, не превышающих 0,5 - 1,5 мм ооцитов. Размеры крупных ооцитов быстро увеличиваются, в связи с чем происходит столь же быстрое истощение жировых тел, как и у самцов. К моменту откладки яиц жировые тела оказываются полностью израсходованными, а затем (к концу июля) их вес снова начинает нарастать.
Помимо изменения жировых тел происходят и связанные с половым циклом изменения в работе гипофиза, гипоталамуса и щитовидной железы (повышенная активность и нейросекреторная деятельность в процессе размножения) (Фелiкс, 1970).
Таким образом, приведенные данные показывают, что развитие половых продуктов у прыткой ящерицы происходит не только за счет вещества жировых тел, но и изменения секреторной деятельности щитовидной железы и гормональной работы гипоталамуса и гипофиза.
Возраст
Точные данные о возрасте наступления половой зрелости у самцов и самок прыткой ящерицы в литературе отсутствуют.
Некоторые авторы (Щепотьев, 1948; Андрушко, 1954; Хонякина, 1964, и др.) предполагают, что половозрелость у прыткой ящерицы наступает в возрасте около двух лет. М. Ф. Тертышников (1972б) определяет наступление половой зрелости для ящериц Ставрополья в 23 месяца. Существует также предположение (Чернов, 1954), что половозрелыми прыткие ящерицы становятся в конце третьего года жизни. Г. П. Лукина (1966б) определяет наступление половой зрелости у самцов и самок в начале третьего года жизни.
Размеры животных
При достижении половой зрелости ящерицы (самцы и самки) имеют вполне определенные размеры (вес, длина). Если о длине тела этой группы можно говорить вполне определенно, то о весовой характеристике половозрелых особей (особенно самок), впервые вступающих в размножение, сказать затруднительно (см. гл. III). Это вызвано тем, что развивающиеся к моменту спаривания яйца искажают действительную весовую характеристику этой группы.
Длина тела. К сожалению литературных данных такого рода очень мало и в некоторых случаях довольно трудно определить какого пола они касаются. Материалы табл. 33 свидетельствуют о том, что размеры самцов в лесостепной зоне несколько больше, чем самок (Жаркова, 1973а). Сведения по степной зоне скудны, а имеющиеся данные не позволяют говорить о превосходстве в размерах одной половой группы над другой. По данным Г. П. Лукиной (1966б), самцы значительно меньше самок, достигших половой зрелости. Самки степной зоны тоже отличаются по своим размерам в разных районах. Они несколько мельче в волжских степях (71,0 ± 0,06, Щепотьев, 1950), чем в более южных районах (Ставрополье и Ростовская обл.), где их длина составляет в среднем 81 - 85 мм (Лукина, 1966б; Тертышников, 1972б). В горных районах (Кутузова, 1971) самки чуть мельче своих сородичей из степной зоны (см. табл. 33). Больше сведений и конкретных данных по другим выделяемым природным зонам (см. гл. V) в литературных источниках нами не обнаружено.
Таблица 33
| Природная зона | Самцы | Самки | ||||
| Lim | M ± m | n | Lim | X ± sx | n | |
| Лесостепная | 61 - 80 | 70,0 - 0,9 | 190 | 55 - 76 | 66,1 ± 0,13 | 169 |
| Степная | 60 - 75 | - | 298 | 76 - 90 | 81,0 ± 0,08 | 23 |
| - | - | - | 60 - 80 | 71,0 ± 0,06 | 65 | |
| - | - | - | 78 - 92 | 85,0 ± 0,01 | 60 | |
| Горные районы | - | - | - | 70 - 85 | - | 15 |
В целом можно заключить, что ящерицы, вступающие в размножение, по мере продвижения от северных границ ареала к южным отличаются по своей длине, и южнее они становятся гораздо крупнее. Видимо исключение составляют горные популяции, которые дают пеструю картину по аналогии с изменением веса в этих районах (см. гл. III).
Вес животных. В литературе не существует данных, которые бы позволили определить весовую категорию, при которой ящерицы достигают половой зрелости. Об этом можно судить лишь по косвенным данным. В главе III были рассмотрены весовая и возрастная структуры популяции, позволившие выделить у самцов весовые классы. Особи, достигшие половой зрелости (возраст был определен по слоистой структуре кости) и впервые вступившие в размножение, распределились в диапазоне от 7 до 11 г. Но и эти данные, по-видимому, не точны, так как здесь играет роль участие в размножении, требующее определенной затраты общей энергии (Щепотьев, 1950). На весовые данные влияют также и климатические условия данного года и связанные с этим условия кормности.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'