Внешний вид и размеры тела
Внешний вид прыткой ящерицы (рис. 33) определяется несколько суженной кпереди плоской головой, длинным и гибким туловищем со сравнительно небольшими, симметрично расположенными передними и задними конечностями и длинным сужающимся хвостом.

Рис. 33. Общий вид взрослых прытких ящериц со спины (А) и с брюшной стороны (Б). Видны основные особенности распределения групп щитков на голове, шее, туловище и основании хвоста (фото А. В. Валецкого)
Голова ящерицы имеет округленно пирамидальную форму. На вершине головы, в центре межтеменного щитка, располагается теменной (третий) глаз - (париетальный орган), значение которого до сих пор не вполне ясно. Глаза располагаются по бокам в средней части головы. Характерной чертой глаз прыткой ящерицы, как и всех рептилий, является хорошее развитие третьего века (мигательной перепонки). Верхние и нижние веки ящерицы подвижны. На конце морды располагаются парные ноздри, а по бокам в задней части головы - вертикальные, овальной формы слуховые отверстия, затянутые в глубине барабанной перепонкой.
Туловище ящерицы удлиненное и гибкое, отделено от головы выраженным шейным перехватом. Границей между туловищем и хвостом является клоака, открывающаяся наружу поперечным отверстием на вентральной стороне тела (см. рис. 33). Хвост в поперечном сечении округлый, постепенно сужающийся к концу. Основание хвоста несколько более массивное у самцов (это связано с расположением крупных парных копулятивных органов позади клоаки).
Хвост ящерицы часто отсутствует в результате аутотомии (отбрасывания) - приспособления, помогающего ящерице избежать преследования хищника. Поэтому по числу особей с аутотомированными животными в популяциях можно косвенно судить о давлении хищников (см. гл. X). При регенерации остающейся части хвоста (или как врожденное уродство) изредка возникает двойной хвост.
Конечности сравнительно небольшие, причем задние развиты заметно сильнее передних. Кисть и стопа оканчиваются пятью тонкими пальцами, вооруженными острыми коготками.
Как линейные размеры, так и вес тела являются признаками комплексными, характеризующимися сравнительно низкими величинами наследуемости; это значит, что они могут рассматриваться в первую очередь в качестве эколого-биогеоценотических показателей. При этом более удобным и точным показателем является общий вес тела, который более объективно, чем любые линейные промеры, может характеризовать место как отдельной особи, так и разных возрастно-половых групп в потоках вещества и энергии, проходящих через населяемые данным видом биогеоценозы.
Вес тела
Данных об изменении веса тела по всему ареалу того или иного вида рептилий мало, и они основаны, как правило, па случайном и неспециально собранном материале (Терентьев, 1946, 1947; Шварц и др., 1968; Lindsey, 1966). Имеющиеся к настоящему времени в нашем распоряжении сведения, включающие Данные по 5423 экз., позволяют проанализировать географические изменения веса тела прыткой ящерицы на большей части ее ареала.
При сопоставлении веса тела ящериц из разных популяций в качестве основного исходного показателя нами был взят максимальный вес самцов. Выбор самцов определен тем, что вес взрослых самок сильно колеблется по месяцам из-за развития яиц и их откладки. Выбор максимального, а не среднего веса определен, во-первых, тем, что, как показал специальный анализ (Смирина, 1974), на средний вес ящериц может оказывать большое влияние количественное соотношение в популяционной выборке разных недифференцируемых при возрасте свыше 2 лет возрастных групп, во-вторых, тем, что не всегда возможно по внешнему виду отличить крупных неполовозрелых особей от мелких половозрелых.
Для удобства анализа полученные данные по максимальному весу самцов были сгруппированы нами в три класса: 8 - 14, 15 - 21, 22 и более грамм. Такое определение данных по весу, безусловно, несколько "огрубляет" их, но вместе с тем дает более четкую картину для рассмотрения изменений этого показателя в масштабах ареала.
При рассмотрении общей картины изменения веса тела прыткой ящерицы (рис. 34) можно проследить определенную тенденцию к увеличению максимального веса особей по мере продвижения от северной границы ареала к южной. Исключением являются районы рек Урала, Дона и северного берега Азовского моря (здесь встречаются ящерицы максимального веса), а также высокогорные районы Закавказья и Заилийского Алатау (где встречаются ящерицы среднего, а не максимального веса).
Причины, обусловливающие различия по весу тела ящериц названных районов от географически близких, можно понять, проведя сравнение близко расположенных одна от другой, но занимающих разные биотопы популяций. Так, выборки из популяций на правом берегу р. Дон в окрестностях г. Верхний Мамон находились на расстоянии 300 - 400 м одна от другой и были отловлены 31 июля - 1 августа 1971 г. Первая выборка взята из прибрежной балки с невысоким полусухим травостоем, редким невысоким кустарником и обилием мест с оголенной почвой. Вторая - в пойменном лесу с густой высокой травой, обилием лещины, ежевики и шиповника. При сравнении весовых характеристик этих выборок оказалось, что взрослые самцы из балки в среднем заметно мельче представителей поймы (5,1 - 15,0 г, а в пойменном лесу 7,1 - 21,0 г). Другой пример касается трех популяций ящериц из района Нарынского залива Бухтарминского водохранилища (Алтайский край). Одна из популяций была отловлена на острове в 600 - 700 м от следующей береговой популяции, третья находилась в 14 км по берегу водохранилища от второй. К моменту нашего исследования период изоляции островной популяции, связанный с затоплением Бухтарминского водохранилища, длился 11 - 12 лет. При сравнении этих трех популяций видно, что животные с острова значительно мельче представителей береговых популяций. Оба примера подтверждают, что популяции, расположенные географически очень близко одна от другой, могут различаться между собой, и эти различия, видимо, связаны с микробиогеографическими (биоценотическими) различиями.
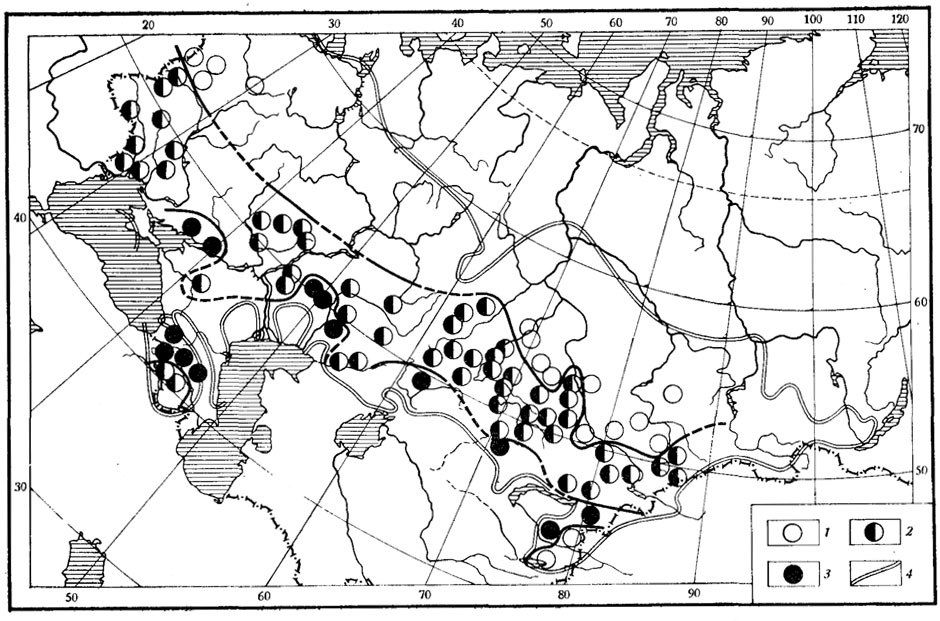
Рис. 34. Географическая изменчивость веса взрослых самцов прыткой ящерицы в центральной и восточной частях ареала. Вес взрослых самцов, в г: 1 - 8 - 14; 2 - 15 - 21; 3 - больше 21; 4 - граница ареала по данным А. С. Баранова и А. В. Валецкого)
В свете только что рассмотренных примеров можно в какой - то степени объяснить и некоторые исключения из общей тенденции изменения веса тела ящериц по ареалу (см. рис. 34), в большинстве случаев относящиеся к горным районам. Последние, как известно, отличаются исключительной пестротой микробиогеографических условий. В подавляющем же большинстве популяций, расположенных на равнинных участках ареала, общее клинальное изменение максимального веса проявляется достаточно отчетливо.
Таблица 3
| Возрастные группы | lim | X | СV* |
| Сеголетки (0 - 3 мес.) | 0,50 - 3,5 | 1,5 | 11 |
| Неполовозрелые (0,5 - 1,5 года) | 2,5 - 6,0 | 4,5 | 22,4 |
| Половозрелые (2 года и старше) | 5,0 - 40,0 | 12,0 | 11 |
*(По данным С. С. Шварца и др. (1968) для Южного Урала.)
В целом полученные данные показывают, что у прыткой ящерицы существуют четко выраженные клинальные изменения максимального веса, прямо противоположные "правилу Бергмана" (Bergman, 1847; Терентьев, 1946). Четко выраженная географическая изменчивость веса указывает на изменение места и значения ящериц в биоценозах разных ландшафтов в пределах всего их огромного ареала.
Как уже говорилось, в отношении самок и молодых животных подобный анализ географической изменчивости веса тела значительно затруднен из-за невозможности точного определения различных возрастных групп при обработке массового материала, высокой степени онтогенетических и сезонных изменений веса. Так, например, недельное голодание приводит к потере до 22, а двухнедельное голодание - до 34% исходного веса взрослыми ящерицами (Шварц и др., 1968). Поэтому мы вынуждены ограничиться здесь приведением лишь общих сводных данных по весу тела основных возрастных и половых групп (см. табл. 3); более подробно данные по весовой динамике роста ящериц рассматриваются в главе VIII (в приложении к биомассе вида и отдельных популяций, в гл. XV).
Данные по резкому увеличению коэффициентов вариации веса тела перезимовавших ящериц указывают на несколько интересных биологических моментов (Шварц и др. 1968). Во-первых, сравнительно невысокая изменчивость молодых перед спячкой свидетельствует об их относительно однородном физиологическом состоянии; резкое увеличение изменчивости вышедших после зимовки ящериц - о разном, видимо, расходе питательных веществ (и энергии) отдельными особями; наконец, в-третьих, снижение изменчивости взрослых вновь свидетельствует о стабилизации основных физиологических отправлений. К сожалению, приведенные данные не позволяют проанализировать изменчивость веса на всем протяжении ареала ящерицы.
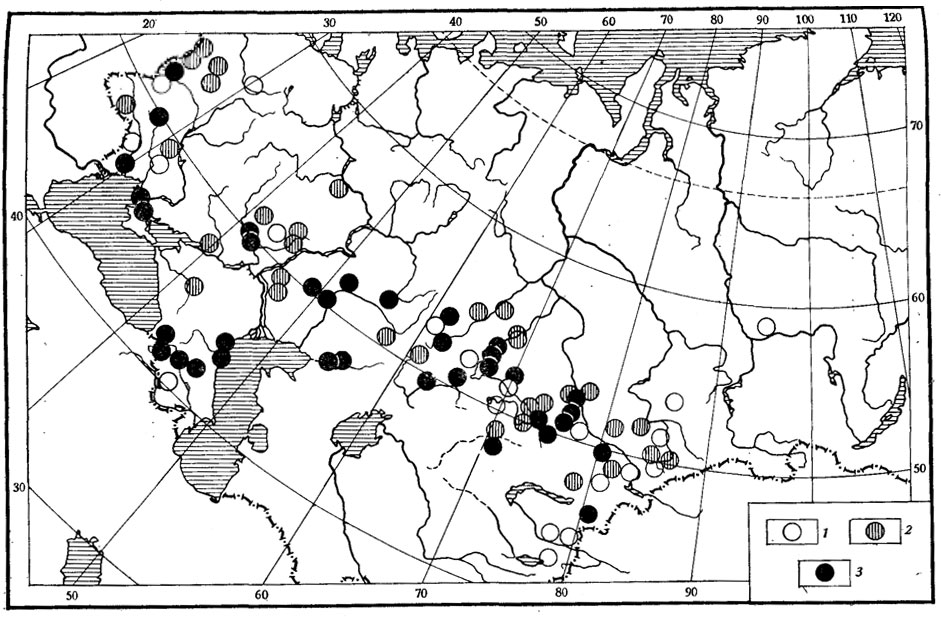
Рис. 35. Географическая изменчивость особенностей полового диморфизма но весу тела прытких ящериц. 1 - самки больше самцов; 2 - половой диморфизм по весу отсутствует; 3 - самки меньше самцов (по данным А. С. Баранова и А. В. Валецкого)
По имеющимся данным, не существует выраженного полового диморфизма по весу тела для вида в целом (рис. 35), поскольку то в одной, то в другой популяции взрослые самки могут быть и больше и значительно меньше, чем самцы.
Длина и другие промеры тела
Таблица 4
| Популяция | Пол | X±Sx | Lim | CV±Scv | n |
| Дагестан | |||||
| Окрестности г. Махачкала | ♂ | 19,92 ± 0,26 | 17,0 - 23,7 | 8,0 ± 0,92 | 38 |
| ♀ | 15,63 ± 0,24 | 14,3 - 18,1 | 7,5 ± 1,08 | 24 | |
| Сергокалинский район | ♂ | 17,23 ± 0,30 | 13,9 - 20,4 | 8,1 ± 1,23 | 22 |
| ♀ | 14,19 ± 0,22 | 12,6 - 17,0 | 7,8 ± 1,08 | 26 | |
| Буйнакский район | ♂ | 17,64 ± 0,30 | 15,5 - 20,0 | 8,8 ± 1,17 | 28 |
| ♀ | 15,35 ± 0,25 | 13,6 - 20,4 | 8,6 ± 1,16 | 27 | |
| Рязанская обл. | |||||
| "Лесостепная популяция" | ♂ | 15,50 ± 0,31 | - | 11,2 ± 1,44 | 33 |
| ♀ | 14,03 ± 0,29 | - | 12,81 ± 1,57 | 33 | |
| "Лесная популяция" | ♂ | 15,36 ± 0,39 | - | 14,7 ± 1,81 | 54 |
| ♀ | 14,00 ± 0,42 | - | 17,4 ± 2,14 | 54 | |
| Горьковская обл. | |||||
| Окрестности с. Старая Пустынь | ♂ | 14,70 ± 0,19 | - | 7,10 ± 0,92 | 30 |
| ♀ | 13,59 ± 0,14 | - | 8,0 ± 0,71 | 63 | |
| Окрестности ст. Сухобезводное | ♂ | 15,03 ± 0,25 | - | 9,3 ± 1,17 | 32 |
| ♀ | 14,16 ± 0,12 | - | 8,1 ± 0,61 | 87 | |
| Армения | |||||
| Севан, островная популяция | ♂ | 15,90 ± 0,30 | - | 12,7 ± 1,69 | 28 |
| ♀ | 13,80 ± 0,44 | - | 14,9 ± 2,30 | 23 | |
| Окрестности с. Семеновки (Севанский район) | ♂ | 13,99 ± 0,40 | - | 9,01 ± 2,02 | 10 |
| ♀ | 12,99 ± 0,38 | - | 13,0 ± 2,1 | 21 | |
Данные по ряду стандартных промеров тела, входящих в таксономическую характеристику данного вида, уже приводились в главе "Систематика". Эти данные свидетельствовали о существовании заметной географической изменчивости многих размерных признаков. Другие промеры тела, характеризующие прытких ящериц разных географических районов, приведены в табл. 4 - 12.
Таблица 5
| Популяция | Пол | X ± Sx | Lim | CV + Scv | n |
| Дагестан | |||||
| Окрестности г. Махачкала | ♂ | 10,18 ± 0,20 | 8,4 - 13,0 | 12,3 ± 1,41 | 38 |
| ♀ | 7,83 ± 0,17 | 7,0 - 10,0 | 10,9 ± 1,57 | 24 | |
| Сергокалинский район | ♂ | 8,45 ± 0,18 | 7,0 - 10,6 | 9,9 ± 1,50 | 22 |
| ♀ | 7,04 ± 0,13 | 6,0 - 8,8 | 9,2 ± 1,28 | 26 | |
| Буйнакский район | ♂ | 8,93 ± 0,24 | 7,0 - 12,3 | 14,3 ± 1,91 | 28 |
| ♀ | 7,44 ± 0,14 | 6,8 - 10,0 | 9,8 ± 1,32 | 27 | |
| Рязанская обл. | |||||
| "Лесостепная популяция" | ♂ | 10,0 ± 0,25 | - | 13,9 ± 1,79 | 33 |
| ♀ | 9,4 ± 0,28 | - | 19,1 ± 2,35 | 33 | |
| "Лесная популяция" | ♂ | 10,06 ± 0,30 | - | 17,6 ± 2,16 | 54 |
| ♀ | 9,41 ± 0,30 | - | 18,4 ± 2,27 | 54 | |
| Горьковская обл. | |||||
| Окрестности с. Старая Пустынь | ♂ | 9,77 ± 0,13 | - | 7,4 ± 0,95 | 30 |
| ♀ | 8,78 ± 0,10 | - | 8,7 ± 0,78 | 63 | |
| Окрестности ст. Сухобезводное | ♂ | 12,69 ± 0,30 | - | 13,4 ± 1,67 | 32 |
| ♀ | 11,51 ± 0,14 | - | 11,2 ± 0,85 | 87 | |
| Армения | |||||
| Севан, островная популяция | ♂ | 10,20 ± 0,20 | - | 12,4 ± 1,65 | 28 |
| ♀ | 9,52 ± 0,22 | - | 11,0 ± 1,70 | 23 | |
| Окрестности с. Семеновки (Севанский район) | ♂ | 10,20 ± 0,41 | - | 13,0 ± 2,91 | 10 |
| ♀ | 9,00 ± 0,26 | - | 13,1 ± 2,07 | 21 | |
Здесь нам кажется целесообразным обратить особое внимание на недостаточную сравнимость приводимых размеров. Это обстоятельство связано с тем, что прыткая ящерица интенсивно растет на протяжении первых лет жизни (см. гл. VIII), и при этом оказывается, что материал, полученный даже в одной и той же популяции в разные месяцы вегетационного сезона, показывает различия. Это относится особенно к младшим и средним возрастным группам. Обычно же авторы суммировали материал, получаемый за все месяцы и за все годы сбора наблюдений. Такие суммарные данные сравнимы только между собой, но не могут дать истинного представления о сезонной и многолетней Динамике размеров тела в тех или иных популяциях, позволяя улавливать лишь самые общие тенденции. В качестве примера анализа таких общих тенденций в размерах тела на рис. 36 приведена длина тела взрослых ящериц из трех разных районов: Крымского полуострова, Предкавказья и Рязанской области. Единственным общим выводом из сопоставления таких данных может быть вывод о существовании во всех этих районах известного полиморфизма по длине тела: более или менее четко выделяются три крупные размерные группы (абсолютные величины, характеризующие каждую из групп особей, везде несколько различны). Как показано ниже (глава "Рост и возраст"), две первые из этих размерных групп более или менее соответствуют ящерицам второго и третьего года жизни, тогда как третья группа, видимо, включает ящериц нескольких старших возрастов. Несомненно, для получения точных сравнимых данных по размерам тела ящериц из разных частей ареала необходимо сравнение ящериц обязательно одного возраста (или, во всяком случае, пойманных в одни и те же достаточно сжатые сроки).
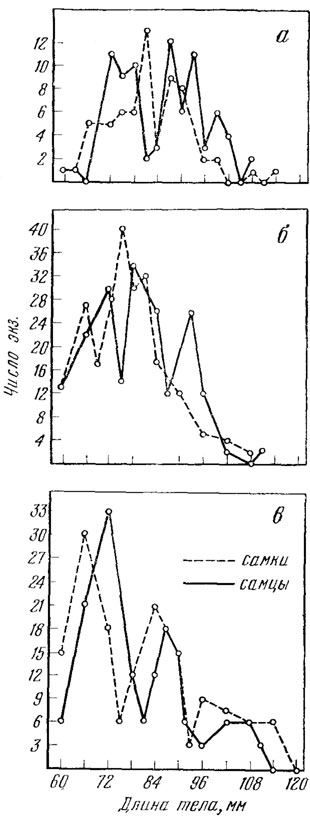
Рис. 36. Распределение самцов и самок (взрослых) по длине тела в трех популяциях. а - Крым (Н. Н. Щербак, 1966); б - Предкавказье (Г. П. Лукина, 1966б); в - Рязанская обл. (В. К. Жаркова, 1973б)
Таблица 6
| Популяция | Пол | X ± Sx | Lim | CV ± Scv | n |
| Дагестан | |||||
| Окрестности г. Махачкала | ♂ | 15,82 ± 0,21 | 13,5 - 18,6 | 8,3 ± 0,95 | 38 |
| ♀ | 12,71 ± 0,22 | 11,0 - 15,5 | 8,3 ± 1,20 | 24 | |
| Сергокалинский район | ♂ | 13,82 ± 0,23 | 11,4 - 16,0 | 7,7 ± 1,17 | 22 |
| ♀ | 11,58 ± 0,19 | 9,7 - 13,7 | 8,4 ± 1,17 | 26 | |
| Буйнакский район | ♂ | 14,14 ± 0,27 | 12,0 - 17,0 | 9,9 ± 1,32 | 28 |
| ♀ | 12,44 ± 0,20 | 11,6 - 16,6 | 8,5 ± 1,15 | 27 | |
| Рязанская область | |||||
| "Лесостепная популяция" | ♂ | 13,26 ± 0,32 | - | 13,3 ± 1,84 | 33 |
| ♀ | 11,66 ± 0,46 | - | 22,8 ± 2,80 | 33 | |
| "Лесная популяция" | ♂ | - | 12,78 ± 0,23 | 10,4 ± 1,28 | 54 |
| ♀ | 11,34 ± 0,36 | - | 18,3 ± 2,24 | 54 | |
| Горьковская область | |||||
| Окрестности с. Старая Пустынь | ♂ | 12,43 ± 0,14 | - | 6,1 ± 0,79 | 30 |
| ♀ | 11,46 ± 0,10 | - | 7,2 ± 0,64 | 63 | |
| Окрестности ст. Сухобезводное | ♂ | 12,06 ± 0,18 | - | 7,5 ± 0,94 | 32 |
| ♀ | 11,44 ± 0,12 | - | 9,4 ± 0,71 | 87 | |
| Армения | |||||
| Севан, островная популяция | ♂ | 13,20 ± 0,20 | - | 8,1 ± 1,08 | 28 |
| ♀ | 11,42 ± 0,33 | - | 13,4 ± 2,06 | 23 | |
| Окрестности с. Семеновки (Севанский район) | ♂ | 10,70 ± 0,44 | - | 13,2 ± 2,96 | 10 |
| ♀ | 10,8 ± 0,35 | - | 14,8 ± 2,33 | 21 | |
Таблица 7
| Популяция | Пол | X + Sx | Lim | CV ± Scv | n |
| Дагестан | |||||
| Окрестности г. Махачкала | ♂ | 93,84 ± 1,01 | 81,0 - 107,8 | 6,6 ± 0,76 | 38 |
| ♀ | 85,08 ± 1,21 | 70,0 - 97,0 | 7,1 ± 1,02 | 24 | |
| Сергокалинский район | ♂ | 84,73 ± 1,35 | 70,5 - 97,0 | 7,5 ± 1,14 | 22 |
| ♀ | 77,38 ± 1,27 | 60,0 - 87,6 | 8,4 ± 1,17 | 26 | |
| Буйнакский район | ♂ | 86,28 ± 1,52 | 70,2 - 106,6 | 9,3 ± 1,24 | 28 |
| ♀ | 82,18 ± 1,10 | 75,4 - 97,0 | 6,9 ± 0,93 | 27 | |
| Рязанская область | |||||
| "Лесостепная популяция" | ♂ | 73,33 ± 1,03 | - | 7,7 ± 0,99 | 33 |
| ♀ | 71,90 ± 1,64 | - | 13,4 ± 1,64 | 33 | |
| "Лесная популяция" | ♂ | 71,44 ± 3,67 | - | 29,5 ± 3,63 | 54 |
| ♀ | 75,42 ± 1,23 | - | 9,4 ± 1,15 | 54 | |
| Горьковская область | |||||
| Окрестности с. Старая Пустынь | ♂ | 75,29 ± 0,74 | - | 5,4 ± 0,70 | 30 |
| ♀ | 74,48 ± 0,76 | - | 8,1 ± 0,72 | 63 | |
| Окрестности ст. Сухобезводное | ♂ | 78,31 ± 1,31 | - | 9,4 ± 1,18 | 32 |
| ♀ | 80,55 ± 0,84 | - | 9,7 ± 0,73 | 87 | |
| Армения | |||||
| Севан, островная популяция | ♂ | 77,00 ± 0,76 | - | 8,0 ± 1,07 | 25 |
| ♀ | 73,71 ± 1,52 | - | 9,4 ± 1,45 | 23 | |
| Окрестности с. Семеновки (Севанский район) | ♂ | 69,00 ± 2,20 | - | 10,2 ± 2,30 | 10 |
| ♀ | 69,10 ± 2,40 | - | 15,8 ± 2,50 | 21 | |
Таблица 8
| Популяция | Пол | X + Sx | Lim | CV ± Scv | n |
| Дагестан | |||||
| Окрестности г. Махачкала | ♂ | 13,11 ± 0,27 | 10,0 - 16,4 | 12,4 ± 1,42 | 38 |
| ♀ | 9,04 ± 0,22 | 7,9 - 11,5 | 11,7 ± 1,69 | 24 | |
| Сергокалинский район | ♂ | 10,18 ± 0,29 | 7,4 - 13,6 | 13,2 ± 2,00 | 22 |
| ♀ | 7,85 ± 0,20 | 6,7 - 10,0 | 13,1 ± 1,82 | 26 | |
| Буйнакский район | ♂ | 10,36 ± 0,28 | 8,0 - 14,3 | 14,5 ± 1,93 | 26 |
| ♀ | 8,33 ± 0,18 | 7,0 - 12,5 | 11,3 ± 1,53 | 27 | |
| Рязанская область | |||||
| "Лесостепная популяция" | ♂ | 10,16 ± 0,33 | - | 18,1 ± 2,33 | 33 |
| ♀ | 8,54 ± 0,22 | - | 15,0 ± 1,84 | 33 | |
| "Лесная популяция" | ♂ | 10,00 ± 0,35 | - | 20,0 ± 2,46 | 54 |
| ♀ | 8,65 ± 0,38 | - | 26,5 ± 1,06 | 54 | |
| Горьковская область | |||||
| Окрестности с. Старая Пустынь | ♂ | 10,77 ± 0,16 | - | 8,2 ± 1,06 | 30 |
| ♀ | 9,70 ± 0,10 | - | 7,9 ± 0,71 | 63 | |
| Окрестности ст. Сухобезводное | ♂ | 11,44 ± 0,23 | - | 11,5 ± 1,44 | 32 |
| ♀ | 10,63 ± 0,12 | - | 10,2 ± 0,77 | 87 | |
| Армения | |||||
| Севан, островная популяция | ♂ | 10,40 ± 0,20 | - | 11,5 ± 1,53 | 28 |
| ♀ | 8,60 ± 0,32 | - | 17,0 ± 2,61 | 23 | |
| Окрестности с. Семеновки (севанский район) | ♂ | 8,79 ± 0,23 | - | 8,5 ± 1,90 | 10 |
| ♀ | 7,89 ± 0,29 | - | 16,9 ± 2,67 | 21 | |
Таблица 9
| Популяция | Пол | X + Sx | Lim | CV ± Scv | n |
| Дагестан | |||||
| Окрестности г. Махачкала | ♂ | 72,24 ± 0,89 | 64,0 - 86,0 | 7,4 ± 0,85 | 33 |
| ♀ | 69,50 ± 1,18 | 57,8 - 80,0 | 8,3 ± 1,20 | 24 | |
| Сергокалинский район | ♂ | 67,91 ± 1,27 | 56,0 - 78,0 | 8,8 ± 1,33 | 22 |
| ♀ | 62,77 ± 1,27 | 48,0 - 72,0 | 10,3 ± 1,43 | 26 | |
| Буйнакский район | ♂ | 68,42 ± 1,06 | 55,0 - 79,6 | 8,2 ± 0,97 | 27 |
| ♀ | 68,11 ± 1,16 | - | 10,4 ± 1,34 | 33 | |
| Рязанская область | |||||
| "Лесостепная популяция" | ♂ | 61,16 ± 1,16 | - | 10,4 ± 1,34 | 33 |
| ♀ | 60,0 ± 1,56 | - | 15,0 ± 1,84 | 33 | |
| "Лесная популяция" | ♂ | 58,62 ± 1,79 | - | 17,6 ± 2,16 | 54 |
| ♀ | 62,78 ± 1,41 | - | 12,8 ± 1,60 | 54 | |
| Горьковская область | |||||
| Окрестности с. Старая Пустынь | ♂ | 60,80 ± 0,63 | - | 5,6 ± 0,73 | 30 |
| ♀ | 61,48 ± 0,69 | - | 8,9 ± 0,79 | 63 | |
| Окрестности ст. Сухобезводное | ♂ | 62,28 ± 1,10 | - | 10,0 ± 1,25 | 32 |
| ♀ | 65,45 ± 0,70 | - | 10,0 ± 0,76 | 87 | |
| Армения | |||||
| Севан, островная популяция | ♂ | 61,00 ± 1,10 | - | 8,4 ± 1,12 | 28 |
| ♀ | 60,57 ± 1,54 | - | 11,7 ± 1,80 | 23 | |
| Окрестности с. Семеновки (Севанский район) | ♂ | 55,19 ± 1,71 | - | 9,8 ± 2,18 | 10 |
| ♀ | 55,80 ± 2,09 | - | 16,7 ± 2,60 | 21 | |
Таблица 10
| Популяция | Пол | X + Sx | Lim | CV ± Scv | n |
| Дагестан | |||||
| Окрестности г. Махачкала | ♂ | 9,29 ± 0,46 | 7,7 - 11,0 | 10,8 ± 1,24 | 38 |
| ♀ | 7,80 ± 0,19 | 6,8 - 10,4 | 11,7 ± 1,69 | 24 | |
| Сергокалинский район | ♂ | 7,91 ± 0,17 | 6,0 - 9,3 | 10,0 ± 1,51 | 22 |
| ♀ | 6,73 ± 0,14 | 5,42 - 8,8 | 10,5 ± 1,46 | 26 | |
| Буйнакский район | ♂ | 8,32 ± 0,19 | 7,4 - 10,8 | 12,0 ± 1,60 | 28 |
| ♀ | 7,33 ± 0,15 | 6,4 - 9,6 | 10,5 ± 1,42 | 27 | |
| Рязанская область | |||||
| "Лесостепная популяция" | ♂ | 7,70±0,20 | - | 14,9±1,92 | 33 |
| ♀ | 6,42 ± 0,49 | - | 25,3 ± 3,10 | 33 | |
| "Лесная популяция" | ♂ | 7,58±0,20 | - | 15,0 ± 1,84 | 54 |
| ♀ | 6,67 ± 0,12 | - | 10,9 ± 1,34 | 54 | |
| Горьковская область | |||||
| Окрестности с. Старая Пустынь | ♂ | 8,40 ± 0,10 | - | 6,5 ± 0,84 | 30 |
| ♀ | 7,81 ± 0,11 | - | 1,11 ± 1,99 | 63 | |
| Окрестности ст. Сухобезводное | ♂ | 8,75 ± 0,18 | - | 11,4 ± 1,43 | 32 |
| ♀ | 8,86 ± 0,12 | - | 12,5 ± 0,94 | 87 | |
| Армения | |||||
| Севан, островная популяция | ♂ | 7,60 ± 0,25 | - | 13,7 ± 1,83 | 28 |
| ♀ | 6,66 ± 0,34 | - | 23,3 ± 3,59 | 23 | |
| Окрестности с. Семеновки (Севанский район) | ♂ | 5,50 ± 0,25 | - | 14,7 ± 3,27 | 10 |
| ♀ | 5,30 ± 0,32 | - | 27,4 ± 4,33 | 21 | |
Таблица 11
| Популяция | Пол | X + Sx | Lim | CV ± Scv | n |
| Дагестан | |||||
| Окрестности г. Махачкала | ♂ | 37,89 ± 0,72 | 33,8 - 47,0 | 11,7 ± 1,34 | 38 |
| ♀ | 34,78 ± 0,50 | 27,0 - 39,7 | 6,8 ± 0,98 | 24 | |
| Сергокалинский район | ♂ | 37,68 ± 0,67 | 31,3 - 41,0 | 8,3 ± 1,26 | 22 |
| ♀ | 32,94 ± 0,40 | 28,0 - 38,8 | 6,3 ± 0,88 | 26 | |
| Буйнакский район | ♂ | 38,07 ± 0,41 | 33,0 - 43,0 | 5,6 ± 0,75 | 23 |
| ♀ | 33,78 ± 0,48 | 28,0 - 40,0 | 7,3 ± 0,99 | 27 | |
| Рязанская область | |||||
| "Лесостепная популяция" | ♂ | 31,2 ± 1,47 | - | 25,9 ± 3,33 | 33 |
| ♀ | 29,67 ± 0,64 | - | 12,5 ± 1,54 | 33 | |
| "Лесная популяция" | ♂ | 30,34 ± 1,54 | - | 28,7 ± 3,53 | 54 |
| ♀ | 29,67 ± 0,64 | - | 12,5 ± 1,54 | 54 | |
| Горьковская область | |||||
| Окрестности с. Старая Пустынь | ♂ | 32,07 ± 0,34 | - | 5,7 ± 0,74 | 30 |
| ♀ | 30,24 ± 0,22 | - | 5,7 ± 0,51 | 63 | |
| Окрестности ст. Сухобезводное | ♂ | 34,92 ± 0,56 | - | 8,1 ± 1,14 | 32 |
| ♀ | 33,38 ± 0,29 | - | 7,7 ± 0,60 | 87 | |
Таблица 12
| Популяция | Пол | X + Sx | Lim | CV ± Scv | n |
| Дагестан | |||||
| Окрестности г. Махачкала | ♂ | 16,89 ± 0,20 | 14,5 - 19,5 | 6,9 ± 0,79 | 38 |
| ♀ | 14,42 ± 0,21 | 11,0 - 16,4 | 7,2 ± 1,06 | 23 | |
| Сергокалинский район | ♂ | 15,68 ± 0,17 | 14,2 - 17,4 | 5,2 ± 0,79 | 22 |
| ♀ | 13,73 ± 0,17 | 12,8 - 16,0 | 6,3 ± 0,88 | 26 | |
| Буйнакский район | ♂ | 16,19 ± 0,16 | 14,8 - 18,0 | 5,3 ± 0,71 | 28 |
| ♀ | 14,67 ± 0,19 | 13,0 - 16,6 | 6,5 ± 0,88 | 27 | |
| Горьковская обл. | |||||
| Окрестности с. Старая Пустынь | ♂ | 12,47 ± 0,13 | - | 5,8 ± 0,75 | 30 |
| ♀ | 11,70 ± 0,11 | - | 7,5 ± 0,67 | 63 | |
| Окрестности ст. Сухобезводное | ♂ | 14,00 ± 0,28 | - | 11,4 ± 1,60 | 32 |
| ♀ | 13,05 ± 0,08 | - | 5,7 ± 0,44 | 87 | |
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'