Таксономия бисексуальных и партеногенетических форм скальных ящериц и современные критерии вида
Выше мы уже говорили, что многие распространенные в предела Кавказа формы скальных ящериц были описаны в свое время как самостоятельные виды и лишь позднее стали рассматриваться специалистам в качестве подвидов одного полиморфного вида Lacerta saxicola Eversmann В связи с тем что многочисленные описанные к настоящему времен: подвиды характеризуются различной степенью морфологической дифференциации и в таксономическом отношении очень неравноценны рядом авторов высказывались предположения, что некоторые из ни: в действительности представляют собой вполне хорошие виды (Mertens 1922; Даревский, 1957; Терентьев, 1961; Добровольская, 1964).
Совершенно очевидно, что дилемма "вид или подвид" в каждом таков случае должна решаться не на основании субъективных мнений тог или иного специалиста, а в первую очередь путем привлечения современных объективных критериев вида. Нужно отметить, что старые авторы, внесшие исключительно большой вклад в дело изучения группы Lacerta saxicola, руководствовались только морфологическими признаками, причем, как правило, не располагали серийным материалом, а работали с единичными особями, происходящими из различных частей ареала. Именно по этой причине Лантц и Цирен (Lantz et Cyren, 1936] отмечали в своей ревизии, что предлагаемая ими схема довольно условна и должна будет совершенствоваться по мере накопления дальнейшие знаний. Неудивительно поэтому, что современные представления о систематике рассматриваемой группы, основанные не только на анализе морфологических признаков, но и на изучении живых ящериц в природе, существенно отличаются от взглядов, которых придерживались прежние исследователи.
Ниже мы постараемся показать, что каждая из рассматриваемых здесь форм скальных ящериц характеризуется определенным комплексом диагностических признаков, позволяющих на основе морфологического критерия достоверно их отличать друг от друга.
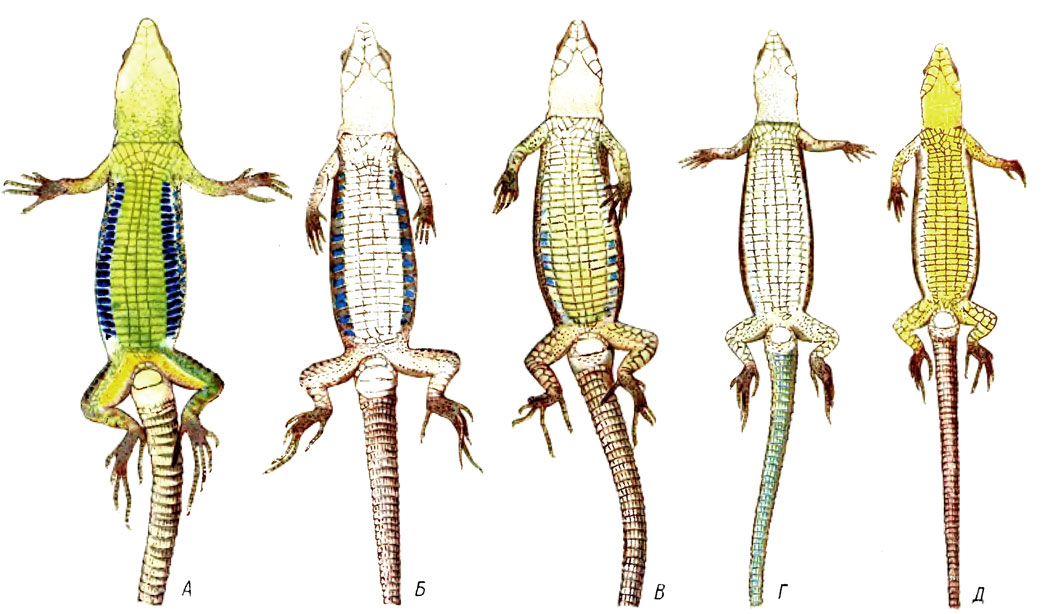
Табл. II Прижизненная окраска нижней стороны тела: А - L. s, nairensis, ♂; Б - L. unisexualis,♀; В - L. armeniaca, ♀; Г - L. dahli, ♀; Д - L. rostombekovi, ♀
Поскольку большинство кавказских представителей этой группы обладает в той или иной мере выраженными перекрывающимися ареалами, для решения вопроса об их таксономическом ранге особое значение при обретает постоянство систематических признаков в зонах совместного обитания двух или нескольких пограничных форм. Иными словами, проблема "вид или подвид" в данном случае должна решаться прежде всего с позиций морфолого-географического критерия вида. Предпринятое нами детальное изучение этого вопроса показало, что во многих случаях морфологические признаки так называемых подвидов при наличии явной географической изменчивости сохраняют свою самостоятельность практически в любой точке ареала, т. е. "подвиды" ведут себя здесь в этом отношении как вполне самостоятельные виды. Последнее обстоятельство особенно интересно, так как, по современным представлениям, при наличии непрерывного ареала подвиды характеризуются обязательным присутствием между ними промежуточных переходных форм, связывающих каждый данный подвид с ближайшим к нему в полосе их соприкосновения. Это положение, хорошо подтверждающееся на многих примерах в орнитологии, было четко сформулировано Э. Майром (Мауг, 1947, 1963), согласно которому "подвиды почти всегда переходят друг в друга незаметно, и в длинной цепи популяций каждая из них представляет собой промежуточную стадию между двумя соседними, соединяя таким образом крайние". Констатированное нами отсутствие промежуточных стадий у ряда кавказских форм Lacerta saxicola, ранее рассматриваемых в качестве отдельных подвидов, является, таким образом, хорошим свидетельством именно в пользу их видовой самостоятельности. Основываясь на этом принципе, мы имеем все основания рассматривать в качестве самостоятельных видов, в частности, такие широко распространенные и морфологически четко различающиеся кавказские формы скальных ящериц, как L. rudis Bedriaga, L. caucasica Mehely и L. saxicola Eversmann, описанные, кстати сказать, в свое время именно как от дельные виды. Характерное для этих видов перекрывание ареалов представлено на рис. 2.
Однако, с другой стороны, анализ географической изменчивости некоторых форм свидетельствует о существовании хорошо выраженных промежуточных зон, образованных особями, сочетающими в себе признаки обоих пограничных подвидов. Иными словами, здесь наблюдается классический случай перехода признаков от подвида к подвиду в полосе соприкосновения их ареалов. Подобные переходные зоны характерны, в частности, для ареалов L. s. darevskii, L. s. brauneri и L. s. saxicola, образующих тесную группу подвидов, распространенных в западной части Большого Кавказского хребта (рис. 1). Таким образом, применение морфолого-географического метода при изучении системы полиморфной группы скальных ящериц, как и следовало ожидать, выявило таксономическую неравноценность многочисленных "подвидовых" форм, среди которых наряду с действительными подвидами оказались и хорошие виды. Отметим также, что если разные виды скальных ящериц отличаются друг от друга прежде всего четко выраженными различиями отдельных элементов чешуйчатого покрова, то различия между подвидами основываются преимущественно на расхождении некоторых меристических показателей фолидоза. Ко всему сказанному можно добавить, что ареалы соседних подвидов скальных ящериц в полосе их соприкосновения, как правило, лишь незначительно перекрывают друг друга, тогда как область совместного обитания симпатрических видов захватывает обычно весьма обширную территорию, где оба они существуют совместно, не обнаруживая каких-либо видимых различий в биотопических требованиях. Нужно отметить, что подобные же зоны совместного обитания двух и даже трех разных видов из близкой к Lacerta saxicola группы европейских стенных ящериц наблюдаются и в ряде мест Южной Европы, причем их видовая самостоятельность в этом случае совершенна не вызывает сомнений (Klemmer, 1957). Перекрывание ареалов L. saxi cola и L. muralis имеет место также в некоторых районах западной Анатолии (Mertens, 1952).
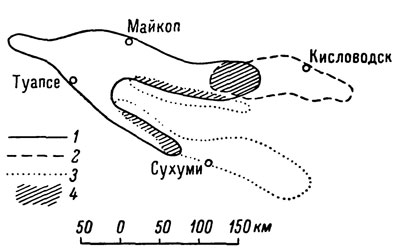
Рис. 1. Схема перекрывания подвидовых ареалов при наличии зоны интерградации: 1 - L. saxicola darevskii; 2 - L. saxicola brauneri; 3 - L. saxicola saxicola; 4 - зона интерградации признаков
Анализ систематических признаков у большого числа особей из смешанных популяций скальных ящериц показывает, что гибриды между ними встречаются сравнительно редко. Это связано с действием биологического механизма репродуктивной изоляции, основанного на несовпадении сроков созревания гонад (стр. 178). В отдельных случаях спаривания самцов и самок бисексуальных форм (о партеногенетических речь будет ниже) получающиеся гибриды обоих полов (F1) всегда плодовиты и способны принимать участие в размножении. Такие гибриды известны, например, между L. s. portschinskii и L. s. raddei и между L. mixta и L. s. parvula. Интересно, что плодовитое потомство получается и при гибридизации скальных ящериц с биологически отличным лесным видом Lacerta derjugini Nik., причем в этом случае гибриды в смешанных по пуляциях встречаются довольно часто.
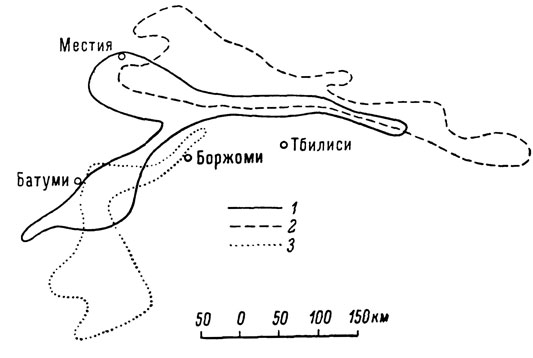
Рис. 2. Схема перекрывания видовых ареалов при отсут ствии зоны ингерградации: 1 - L. rudis rudis; 2 - L. caucasica caucasica; 3 - L. saxicola parvula
В качестве дополнительного критерия, играющего роль при разграничении систематически близких подвидов скальных ящериц, в ряде случаев могут служить и некоторые этологические особенности, в частности манера спаривания и особенности брачного поведения (стр. 174), Как известно, определенную роль при систематических построениях и выяснении родственных отношений животных может играть хромосомный анализ. В отношении группы Archaeolacerta первые исследования такого рода были проведены В. Н. Куликовой в Институте цитологии АН СССР. Куликова показала, что у изученных ею видов и подвидов (L. s. nairensis, L. s. portschinskii, L. s. parvula, L. s. valentini, L. armeniacay L. dahli, L. rostombekovi, L. r. rudis, L. r. obscura, L. mixta и L. derjugini) диплоидное число хромосом равно 38. Можно отметить, что, по данным Матте (Matthey, 1949), аналогичное диплоидное число хромосом установлено у европейской Lacerta muralis и ряда других видов сем. Lacertidae.
Совершенно особую проблему представляет собой вопрос о таксономическом ранге партеногенетических форм скальных ящериц, поскольку применение к ним рассмотренных выше видовых критериев наталкивается на серьезные трудности.
Как мы увидим дальше, область распространения партеногенетических форм armeniaca, dahli, rostombekovi, а также партеногенетической расы обоеполого подвида L. s. nairensis занимает обширную территорию в пределах северной Армении, южной Грузии, северо-западного Азербайджана и северо-восточной Турции, где все они обладают в значительной мере перекрывающимися ареалами (рис. 3). До последнего времени ящерицы эти, как и все бисексуальные формы L. saxicola, рассматривались специалистами в качестве отдельных, четко различающихся подвидов. Достоверность различий между ними, заключающихся прежде всего в особенностях чешуйчатого покрова, была подтверждена недавно методами биометрического анализа (Добровольская, 1964). В целом различия эти таковы, что, основываясь только на морфологическом критерии, мы имеем все основания рассматривать партеногенетические формы в качестве вполне самостоятельных видов. Однако достигнутая к настоящему времени высокая степень изученности этих ящериц позволяет привлечь для решения вопроса об их таксономическом ранге также географический и физиологический критерии вида. Как уже говорилось, все известные партеногенетические формы Lacerta saxicola распространены главным образом в пределах восточного Закавказья, где их ареалы почти не выходят за границы лесной и горно-степной частей Закавказского нагорья в зоне 900-2200 м над ур. м.
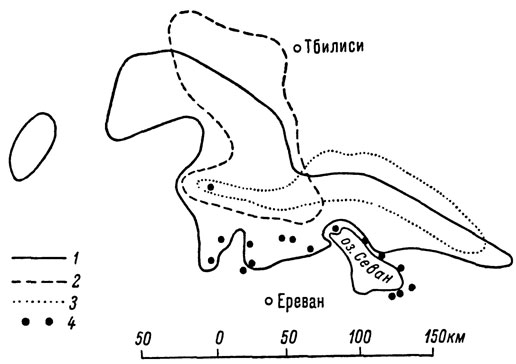
Рис. 3. Схема перекрывания ареалов партеногенетических видов при отсутствии зон интерградации: 1 - L. armeniaca; 2 - L. dahli; 3 - L. rostombekovi; 4 - место нахождения L. unisexualis
Представление об этом дают карты (рис. 26, 32, 35, 53), на которые нанесены основные пункты достоверных находок каждой из четырех рас сматриваемых форм в отдельности. Первое, что обращает на себя внимание при анализе этих ареалов, это их тесная общность. Как видно на рис. 3, ареалы dahli и rostombekovi на значительном протяжении укладываются внутри ареала более широко распространенной формы armeniaca, с которым на юге совпадает также и область распространения партеногенетической расы L. s. nairensis. К этому необходимо добавить, что, обитая в со вершенно сходных экологических условиях, партеногенетические ящерицы не образуют каких-либо промежуточных форм, т. е. ведут себя в этом отношении как самостоятельные виды. Во многих районах север ной Армении и Южной Грузии можно наблюдать, как особи двух или даже трех различных "подвидов" держатся буквально бок о бок, греются рядом на солнце и скрываются при опасности в одно убежище.
Иными словами, партеногенетические ящерицы совершенно не отвечают основному требованию географического критерия, говорящему о не совместимости в общем ареале двух или нескольких подвидов одного вида. Таким образом, применяя для установления таксономического ранга партеногенетических форм Lacerta saxicola морфолого-географический метод, мы имеем все основания считать их самостоятельными видами. Данные, которыми мы располагаем в настоящее время, позволяют привлечь для решения этого вопроса также и физиологический или генетический критерий вида, основанный, как известно, на том факте, что помеси между различными видами в отличие от таковых между подвидами, как правило, бывают бесплодны. Отсутствие собственных самцов исключает гибридизацию между обитающими совместно партеногенетическими формами скальных ящериц, однако на границах ареалов партеногенетические самки получают возможность спариваться с преследующими их самцами сосед них обоеполых подвидов Lacerta saxicola, в результате чего между ними возникают гибриды. Такие гибридные особи всегда являются стерильными триплоидными (3n=57) самками, бесплодие которых обусловлено глубокими аномалиями в развитии и строении гонад (Даревский и Куликова, 1964). Таким образом, физиологический критерий вида также указывает на видовую самостоятельность партеногенетических форм. Подобное решение вопроса действительно было бы возможно в случае, если бы мы имели дело с видами, слагающимися из самцов и самок и размножающимися обычным половым путем. Однако тот факт, что партеногенетические самки не нуждаются в оплодотворении, вынуждает нас рассмотреть вопрос, возможно ли вообще ставить знак равенства между нормальными обоеполыми видами и формами, размножающимися без участия самцов. Вопрос этот далеко не нов и тесно связан с такой важной и по сути дела еще не решенной проблемой, как определение вида. Известно, что современное, так называемое биологическое определение вида основывается на критерии фактической или потенциальной способности к скрещиванию естественных популяций, внутри которых свободно осуществляется панмиксия. Такое определение в качестве непременного условия предполагает существование в популяциях обоих полов и, следовательно, совершенно не приемлемо при отсутствии настоящего полового размножения, частным случаем которого является партеногенез. Так, специально занимавшийся изучением этого вопроса Т. Добржанский полагает даже, что "у бесполых организмов и организмов с облигатным самооплодотворением вид, как наиболее постоянная и поэтому менее произвольная по сравнению с другими категория, отсутствует" (цит. по: Кэйн, 1958). Учитывая очевидную ограниченность биологического определения, некоторые авторы предложили типологию видов, основанную на различных способах их размножения. Одна из наиболее успешных, на наш взгляд, попыток такого рода принадлежит А. Кэйну (1958), по мнению которого необходимо четко отграничивать понятие биологического вида, отличая его от случаев, когда оно неприложимо из-за отсутствия настоящего полового размножения. Такие формы Кэйн для удобства предлагает называть "агамными видами", полагая в то же время, что критерии их видовой самостоятельности должны быть такими же, как у обычных обоеполых форм, и в системе их следует помещать рядом с их ближайшими родичами.
Целесообразность выделения агамных, в том числе и партеногенетических, форм на правах самостоятельных видов, если к этому имеются морфологические предпосылки, была поддержана также Симпсоном (Simpson, 1961) и Майром (Мауг, 1963), хотя, по мнению Н. В. Тимофеева-Ресовского (1965), попытки отождествления видов и подвидов у агамных и бисексуальных форм имеют лишь условный характер. Признавая таксономическую равноценность биологических и агамных видов, мы тем самым положительно решаем вопрос и о возможности применения к ним критериев, основанных на морфологических признаках, поскольку с этой точки зрения принципиальных различий между первыми и вторыми не существует. Иначе, однако, обстоит дело с применением к агамным формам географического и физиологического критериев вида, уже само определение которых тесно связано с наличием настоящего полового размножения. Действительно, допуская существование в границах одного ареала двух или нескольких четко разграниченных видов, мы тем самым предполагаем существование у них особых изолирующих механизмов, препятствующих перекрестному скрещиванию. Механизмы эти, как известно, бывают весьма различны, однако во всех случаях они направлены на недопущение продуктивного спаривания "своих" самцов с самками чужого вида. Совершенно очевидно, что при практическом отсутствии самцов, как это наблюдается в нашем случае, необходимость в возникновении специальных изолирующих механизмов отпадает, ибо каждая отдельная самка с самого начала репродуктивно уже изолирована от всех остальных особей в популяции. По этой причине даже само понятие "популяция", как замечает Майр (Мауг, 1963), едва ли применимо по отношению к такой общности репродуктивно изолированных партеногенетических самок.
Высказанные соображения в полной мере относятся и к случаям применения в отношении партеногенетических форм физиологического критерия вида, основанного на действии цитолого-генетических механизмов, препятствующих получению плодовитых межвидовых гибридов. Так, мы показали (Даревский и Куликова, 1964), что бесплодие гибридов между партеногенетическими и бисексуальными формами скальной ящерицы связано именно с наличием однополого размножения у материнской формы и, следовательно, не должно приравниваться к случаям, когда стерильное потомство возникает в результате гибридизации нормальных биологических видов.
Необходимо, таким образом, выяснить, возможно ли в одном ряду с изолирующими механизмами, действие которых основано, например, на особенностях строения гениталий, несовпадении сроков созревания гонад, специфике брачного поведения, понижения эффективности спаривания и т. д., рассматривать также и нормальное отсутствие самцов, уже само по себе исключающее всякую возможность перекрестного оплодотворения. Отсюда закономерно вытекает и второй важный вопрос, должен ли систематик при оценке таксономического ранга изучаемых им форм принимать во внимание характер механизма репродуктивной изоляции или же он может довольствоваться лишь чисто внешней стороной дела, т. е. уже самим фактом существования такой изоляции. Думается, что выше мы уже принципиально ответили на эти вопросы, когда согласились с необходимостью разделения агамных и биологических видов, основываясь на способе размножения. Точно так же, как, признавая равноценность этих категорий перед лицом таксономии, систематик в то же время не закрывает глаза на принципиальные их различия в биологическом смысле, так и в своей практической работе он не должен делать различий и между механизмами, обеспечивающими репродуктивную изоляцию, поскольку в каждом случае с их помощью обеспечивается важная для него таксономическая самостоятельность изучаемых форм. Применяя это положение к нашему случаю с партеногенетическими ящерицами, необходимо признать, что все они являются не подвидами, а вполне самостоятельными агамными видами, в соответствии с чем и должны именоваться Lacerta armeniaca Mehely, L. dahli Dar., L. rostombekovi Dar. Так же обстоит дело и с нартеногенетической расой бисексуальной формы L. saxicola nairen sis, которая, как мы ранее показали (Даревский, 1962), обладает достоверными морфологическими отличиями и хорошо выраженным ареалом.
Для сравнения отметим, что видовым статусом обладают также много численные установленные в последнее время формы американских партено-генетических ящериц-тейид рода Cnemidophorus, некоторые из которых рассматриваются в качестве видов-двойников (Wright, 1966; Low а. Wright, 1966a, 1966b).
Интересно, что рассмотренный нами случай со скальными ящерицами чрезвычайно напоминает разбираемый А. Кэйном (1958) пример с партено генетическими расами широко распространенного в северном полушарии дождевого червя (Eiseniella tetraedra). Специалистам известны по крайней мере 4 хорошо различающиеся симпатрические формы этого вида, которые с полным правом считают самостоятельными видами, хотя они представ лены исключительно самками и, следовательно, лишены возможности перекрестного оплодотворения. Несомненно, что подобные же случаи могут быть обнаружены и среди многих других партеногенетически размножающихся животных.
Мы уже говорили, что во многих отношениях скальные ящерицы Кавказа обнаруживают значительную аналогию с группой европейских стенных ящериц, относящихся к общему для тех и других подроду Archaeo lacerta. Согласно последним сводкам (Klemmer, 1957; Mertens u. Wermuth, 1960), в Европе этот подрод представлен 6 хорошо различающимися видами с 10 подвидами, общий ареал которых спорадически простирается от центральной и северной Испании на западе до северо-западной части Балканского полуострова и южной Греции на востоке. Морфологические признаки, по которым различаются между собой все европейские и кавказские виды этого подрода, по своей значимости абсолютно не уступают друг другу, и разделение каждой из этих групп на несколько самостоятельных видов с точки зрения систематика вполне оправдано.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'