Б. Односторонняя экстирпация обонятельной плакоды
Для выяснения вопроса о связи закладки собственно хоаны с органом обоняния и материале, участвующем в ее образовании, была поставлена вторая серия опытов, заключавшаяся в односторонней экстирпации обонятельной плакоды. Вторая сторона оперированного экземпляра служила контролем.
Бесхвостые амфибии. Операции производились на эмбрионах травяной и зеленой лягушек, чесночницы и обыкновенной жерлянки на стадиях ранней, средней и поздней хвостовой почки, а также у молодых личинок с только что появившимися жаберными бугорками. Видовая принадлежность так же, как в первой серии опытов у бесхвостых амфибий, не оказала влияния на полученные результаты, и поэтому подопытный материал рассматривается в совокупности. Всего было прооперировано 104 экз., из них выжило 94. Фиксация материала производилась в те же сроки, обусловленные теми же соображениями, что и в серии с трансплантацией обонятельной плакоды у бесхвостых амфибий, за исключением личинок обыкновенной жерлянки, часть которых выращивалась до предметаморфозного периода с целью выяснения возможных преобразований заложившейся хоаны. Окраска препаратов производилась тотально борным кармином и на стеклах квасцовым гематоксилином и по Мал лори.
Из 94 исследованных личинок у 53 орган обоняния отсутствовал полностью, у 18 образовался его регенерат в виде комка клеток, пузырька или тяжа, отходящего от покровов и не имеющего никакой связи с ротовой полостью, и у 23 развился более или менее полноценный орган обоняния, открывающийся в ротовую полость нормальной хоаной.1 Естественно, что последняя группа подопытных личинок исключается из рассматриваемого материала, а первые две можно объединить в одну, состоящую из 71 экз., поскольку при исследовании препаратов выяснилось, что никакой разницы в отношении образования хоаны между личинками первой и второй групп не наблюдается, а следовательно, наличие регенерата органа обоняния, не имеющего связи с ротовой полостью, не оказывает влияния на возникновение хоаны и степень ее развития. Результаты оказались следующими.
1 (Поскольку разбираемые в настоящей работе вопросы не касаются проблемы регенерации, нет смысла приводить более подробные сведения, но все же необходимо упомянуть, что в этом отношении наблюдалась разница между отдельными видами и наибольшую способность к регенерации органа обоняния показали зеленая лягушка и чесночница.)
1. Орган обоняния отсутствует, хоаны нет или наличие ее сомнительно, так как на том месте, где она должна находиться, имеется только небольшое углубление безо всяких следов хоанальных клапанов - 14 случаев.
2. Имеется регенерат органа обоняния, не соединенный с ротовой полостью, хоаны нет или ее наличие сомнительно - 8 случаев.
3. Орган обоняния отсутствует, образовалась нормальная хоана с двумя хоанальными клапанами (рис. 38, А) - 4 случая.
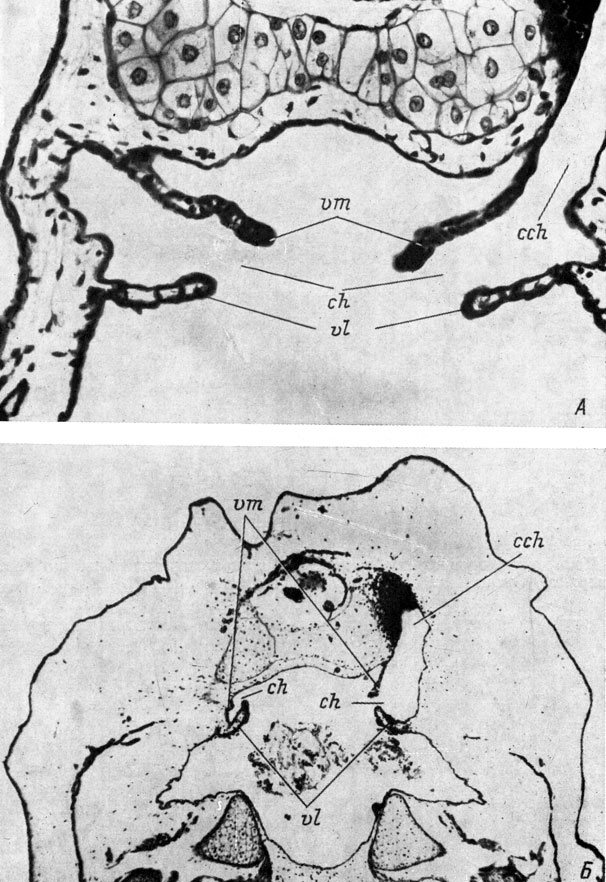
Рис. 38. Поперечные срезы через область хоаны у оперированных личинок Bombina bombina длиной 13 мм (А - нормальная хоана, увел. 190) и 20 мм (Б - дефектная хоана, увел. 40). Слева орган обоняния отсутствует
4. Имеется регенерат органа обоняния, не соединенный с ротовой полостью, образовалась нормальная хоана - 1 случай.
5. Орган обоняния отсутствует, образовалась дефектная, зачастую уменьшенная, хоана с недоразвитием одного или обоих хоанальных клапанов (рис. 38, Б) - 35 случаев.
6. Имеется регенерат органа обоняния, не соединенный с ротовой полостью, образовалась дефектная хоана - 9 случаев.
Для наглядности сопоставления результатов удаления обонятельной плакоды у бесхвостых амфибий полученные данные сведены в табл. 1.

Таблица 1. Зависимость образования хоаны от наличия органа обоняния у бесхвостых амфибий
Из приведенных результатов видно, что наличие регенерата органа обоняния, не соединенного с ротовой полостью, не оказывает никакого влияния на развитие хоаны и при рассмотрении их выделенные группы личинок могут быть объединены попарно: первая со второй, третья с четвертой и пятая с шестой. Оценивая результаты, полученные в первой и второй группах, необходимо отметить, что все относящиеся к этим группам личинки были зафиксированы на довольно ранних стадиях развития, и к моменту фиксации на неоперированной стороне либо еще не оказалось сквозного соединения полости органа обоняния с ротовой полостью, либо оно наступило недавно, поэтому отсутствие оформленной хоаны можно отнести за счет запаздывания ее развития на оперированной стороне. Таким образом, эти данные не противоречат полученным у остальных подопытных личинок, у которых отмечено морфологически независимое от органа обоняния возникновение хоаны как при его полном отсутствии, так и при наличии регенерата, не соединенного с ротовой полостью.
Третья и четвертая группы личинок насчитывают 5 случаев с образованием совершенно нормальной хоаны, причем 2 таких случая наблюдались у обыкновенной жерлянки, 1 у чесночницы и 2 у зеленой лягушки, что означает одинаковую возможность развития нормальной хоаны как у высших, так и у более примитивных бесхвостых амфибий. В трех из этих случаев, кроме самой хоаны, развилась также и проксимальная, прилегающая к хоане, часть хоанального канала (2 случая у обыкновенной жерлянки и 1 у зеленой лягушки).
Пятая и шестая группы личинок включают в себя подавляющее число случаев, а именно 44, с образованием дефектной хоаны. Ее дефектность выражается в меньшей величине углубления и в недоразвитии одного, а иногда и обоих хоанальных клапанов. Возраст, до которого выращивались личинки, не играет роли в развитии нормальной или дефектной хоаны, так как и при фиксации в предметаморфозном периоде хоана оставалась дефектной. В 2 случаях образование дефектной хоаны сопровождалось одновременным развитием проксимальной части хоанального канала (1 случай у травяной и 1 у зеленой лягушки).
Таким образом, закономерным результатом удаления органа обоняния является возникновение хоаны при его полном отсутствии и безо всякой морфологической связи с ним. Следовательно, хоана бесхвостых амфибий развивается самостоятельно и не из ткани обонятельного мешка, а из материала стенки кишечной трубки, а именно той ее части, которая представляет собой ротовую полость. В ином случае образование хоаны в отсутствие органа обоняния (рис. 38) было бы невозможным. Отсюда нельзя говорить о прорыве полости хоанального канала в ротовую полость, как это обычно принято, так как в данном случае имеет место вторичное соединение двух морфологически независимо возникающих образований: собственно хоаны как производного либо эктодермальной стенки ротового впячивания (чесночница), либо энтодермальной стенки передней кишки (обыкновенная жерлянка, травяная и зеленая лягушки) с хоанальным выростом обонятельного мешка. Этот факт полностью согласуется с изложенными выше данными по нормальному развитию хоанального канала у бесхвостых амфибий (глава II), показывающими, что образование хоаны начинается до соединения хоанального выроста обонятельного мешка со стенкой кишечной трубки.
Морфологическая независимость закладки хоаны от органа обоняния у бесхвостых амфибий отнюдь не означает полной независимости ее развития. Поскольку при отсутствии органа обоняния в подавляющем большинстве случаев образуется не полноценная, а дефектная несовершенная хоана, можно думать о наличии индукционного формативного воздействия на нее со стороны этого органа. На первый взгляд этому заключению противоречит, во-первых, отсутствие такого воздействия на развитие хоаны не связанного с ротовой полостью регенерата органа обоняния и, во-вторых, образование хотя бы и в небольшом числе случаев совершенно нормальной хоаны. Однако именно возможность развития в виде исключения нормальной хоаны с обоими клапанами, а иногда и с проксимальной частью хоанального канала при полном отсутствии органа обоняния показывает, что причиной возникновения неполноценной, дефектной, хоаны, наблюдаемого в большинстве случаев у оперированных личинок, является не недостаток материала отсутствующего обонятельного мешка, который может частично, как я допускала раньше (Медведева, 1960а), участвовать в построении хоаны, а отсутствие необходимого формативного воздействия, поскольку хоана способна образоваться исключительно из материала стенки кишечной трубки. Все же механизм этого индукционного взаимодействия между органом обоняния и хоаной у бесхвостых амфибий еще явно несовершенен и не установился полностью. Об этом, во-первых, говорит сам факт возникновения хоаны после удаления закладки органа обоняния, во-вторых, то, что наличие не связанного с ротовой полостью регенерата не оказывает влияния на степень ее развития, и, в-третьих, возможность образования в отдельных случаях совершенно нормальной хоаны. Последнее обстоятельство служит также доказательством исходной независимости развития хоаны и сравнительно недавнего возникновения формативной взаимосвязи с органом обоняния. Допущение возможности предварительной детерминации места закладки хоаны в результате более длительного индуктивного воздействия еще не удаленной обонятельной плакоды исключается, так как образование совершенно нормальной хоаны имело место при операциях на более ранних стадиях развития, чем те, при операции на которых возникала дефектная хоана. Кроме того, наличие случаев с образованием не только самой хоаны, но и каудальной части хоанального канала при отсутствии органа обоняния указывает на то, что исходно материал ротовой полости у бесхвостых амфибий в большей мере участвовал в построении хоанального канала, как это наблюдается в настоящее время у хвостатых амфибий, и подтверждает высказанное выше предположение (см. стр. 48) об усилении роста у бесхвостых амфибий хоанального выроста обонятельного мешка и замещении им выроста передней кишки.
Хвостатые амфибии. При односторонней экстирпации обонятельной плакоды у хвостатых амфибий операции производились на стадиях 21 - 30 по Глезнеру у обыкновенного тритона, на соответствующих им стадиях у кавказского подвида гребенчатого тритона, Ranodon sibiricus и Hynobius keyserlingii и на стадиях 24 - 30 по Гальену и Дюроше у испанского тритона. В этой серии опытов личинки выращивались значительно дольше, чем в серии с трансплантацией обонятельной плакоды, а именно до конца метаморфоза. Для выяснения хода образования хоаны в эксперименте проводилась и промежуточная фиксация на более ранних стадиях развития. Срезы окрашивались по Маллори. Всего было выращено, зафиксировано и исследовано микроскопически 239 экз., из них у 54 личинок имела место регенерация органа обоняния и установилась его связь с ротовой полостью путем образования хоанального канала, поэтому эти личинки в рассматриваемый материал не входят. Следует отметить, что образование достаточно полноценного регенерата наблюдалось у личинок, оперированных на стадии поздней нейрулы и ранней хвостовой почки, а также в некоторых случаях - на стадии средней хвостовой почки. У личинок, оперированных на более поздних стадиях, были только отдельные случаи частичной регенерации, по-видимому за счет неполного удаления плакоды. Из оставшихся 185 личинок у 151 орган обоняния полностью отсутствовал, у 25 наблюдалась частичная регенерация органа обоняния, без соединения его с ротовой полостью, и у 9 образовался слезно-носовой проток, в трех случаях впадающий в хоану. В табл. 2 показано распределение личинок различных видов по приведенным выше результатам операций.
Поскольку образование хоаны имело место во всех трех выделенных группах, в том числе и в той, в которой орган обоняния полностью отсутствует, можно с уверенностью считать, что оно, как и у бесхвостых амфибий, не зависит от наличия не соединенного с ротовой полостью регенерата обонятельного мешка или слезно-носового протока, поэтому весь материал рассматривался вместе. Так как закладка слезно-носового протока не имеет отношения к органу обоняния (И. И. Шмальгаузен, 1958г; Медведева, 1959, 1960б, 1960в), было сочтено возможным присоединить 3 случая со впадением слезно-носового протока в хоану к остальному материалу. Весь рассматриваемый материал можно разделить по результатам в отношении образования хоаны на следующие группы.
1. Никаких следов образования хоаны не имеется - 45 случаев.
2. Образование хоаны сомнительно (на ее месте есть небольшое углубление) - 27 случаев.
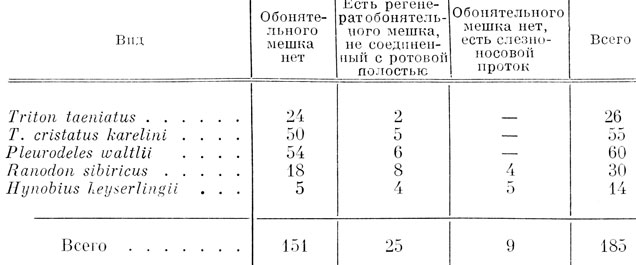
Таблица 2. Результаты удаления обонятельной плакоды у хвостатых амфибий
3. Имеется хорошо развитая хоана, но без клапана - 94 случая.
4. Образовалась совершенно нормальная хоана с медиальным клапаном - 19 случаев.
В первой группе, с полным отсутствием хоаны, 17 случаев относятся к фиксации личинок на довольно ранних стадиях развития, когда хоана на неоперированной стороне еще не образовалась. 21 случай относится к фиксации личинок вскоре после образования хоаны в норме, т. е. не старше, чем стадия 45 по Глюкзон у обыкновенного тритона, стадия 47 по Глюкзон у кавказского подвида гребенчатого тритона и стадия 40 по Гальену и Дюроше у испанского тритона. Ни Ranodon sibiricus, ни Hynobius keyserlingii в эту подгруппу не входят. Поскольку у высших тритонов, как уже отмечалось в описании нормального развития, окончательное образование хоаны, включая клапан, происходит значительно позже, чем у Hynobiidae, можно полагать, что в условиях эксперимента оно еще дополнительно замедляется, и тогда все эти случаи следует отнести к предыдущей подгруппе, когда хоана по времени еще не успела образоваться. И наконец, оставшиеся 7 случаев падают на период метаморфоза, когда на неоперированной стороне исследованных личинок хоанальный клапан либо уже исчез, либо значительно уменьшился по сравнению с нормой, т. е. область хоаны охвачена процессом резорбции. Опять-таки в этой подгруппе нет ни одного представителя ни Ranodon sibiricus, ни Hynobius keyserlingii. Очевидно, у высших тритонов в отсутствие органа обоняния во время метаморфоза не только резорбируется хоанальный клапан, как в норме, но и захваченная процессом резорбции исчезает целиком вся хоана.
Все 27 случаев второй группы, в которой наличие хоаны сомнительно, по времени фиксации примерно совпадают со второй подгруппой первой группы, т. е. фиксация производилась вскоре после образования хоаны на неоперированной стороне. Поэтому с полным основанием можно считать, что в эту группу попали все случаи начального образования хоаны, которое на оперированной стороне запаздывает по сравнению с нормой, В этой группе также не имеется ни одного представителя сем. Hynobiidae.
Таким образом, совершенно очевидно, что все имеющиеся отрицательные и сомнительные случаи не противоречат остальному материалу, показывающему наличие образования хоаны у хвостатых амфибий при полном отсутствии органа обоняния, так же как это было продемонстрировано соответствующими опытами у бесхвостых амфибий.
Самая большая по количеству экземпляров третья группа, с образованием хоаны без клапана, по своему составу также не однородна. Сюда относятся, во-первых, 69 случаев образования дефектной хоаны без клапана вообще или же с очень маленьким выступом эпителия, но не на медиальной стенке хоаны, где клапан образуется в норме, а на ее латеральной стенке, когда на неоперированной стороне имеется нормальный хоанальный клапан (рис. 39, А, Б). Вышеупомянутый выступ эпителия (рис. 39, Б) нельзя считать недоразвившимся клапаном не только потому, что он располагается на латеральной, а не на медиальной стенке хоаны, но также и вследствие образования его в виде незначительного выступа, а не длинной складки, как нормальный хоанальный клапан. Все случаи этой подгруппы опять-таки касаются только обыкновенного, кавказского подвида гребенчатого и испанского тритонов. Ни Ranodon sibiricus, ни Hynobius keyserlingii в эту подгруппу не входят, а составляют целиком следующую подгруппу третьей группы, не содержащую представителей трех вышеупомянутых видов и включающую в себя 23 случая, когда на неоперированной стороне хоанальный клапан еще не образовался или только начал образовываться (фиксация, близкая по срокам к моменту образования хоанального клапана в норме). Такую хоану нельзя назвать дефектной, она только несколько отстает по степени развития клапана от хоаны неоперированной стороны. Эта подгруппа вполне заслуживает включения в четвертую группу с образованием нормальной хоаны. И, наконец, имеются еще два случая (оба у личинок H. keyserlingii, зафиксированных в период метаморфоза) с отсутствием хоанального клапана как на оперированной, так и на неоперированной стороне, хоана же развита на оперированной стороне вполне нормально. Следовательно, у Hynobiidae во время метаморфоза на оперированной стороне резорбируется только клапан, так же как и в норме, хоана же сохраняется, несмотря на полное отсутствие органа обоняния, а не исчезает, как это имеет место у высших хвостатых амфибий. Эти два случая, конечно, тоже должны быть отнесены к четвертой группе с образованием нормальной хоаны.
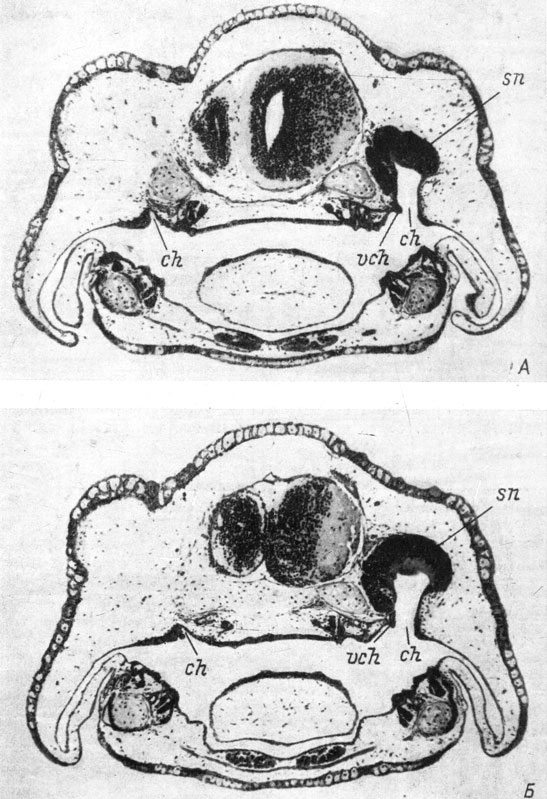
Рис. 39. Поперечные срезы через область хоаны у оперированных личинок Pleurodeles waltlii длиной 22.5 мм, стадия 54 (А - хоана без клапана) и длиной 22 мм, стадия 54 (Б - на латеральной стенке хоаны есть маленький вырост эпителия). Слева орган обоняния отсутствует. (Увел. 38)
Последняя, четвертая, группа с образованием совершенно нормальной хоаны с клапаном, ничем не отличающейся от хоаны неоперированной стороны, включает только личинок Ranodon sibiricus и Hynobius keyserlingii, зафиксированных до наступления метаморфоза, когда хоаны на обеих сторонах развиты одинаково и обладают нормальными хоанальными клапанами (рис. 40). Общее число случаев с образованием хоаны составляет, таким образом, 113 из 185, причем полностью нормальная хоана развилась в 44 случаях (только сем. Hynobiidae - объединены случаи четвертой группы со случаями двух последних подгрупп третьей группы), а дефектная хоана без клапана - в 69 случаях (только сем. Salamandridae). Отсутствие хоаны в остальных случаях объясняется преждевременной фиксацией личинок или же редукцией хоаны во время метаморфоза у Salamandridae.
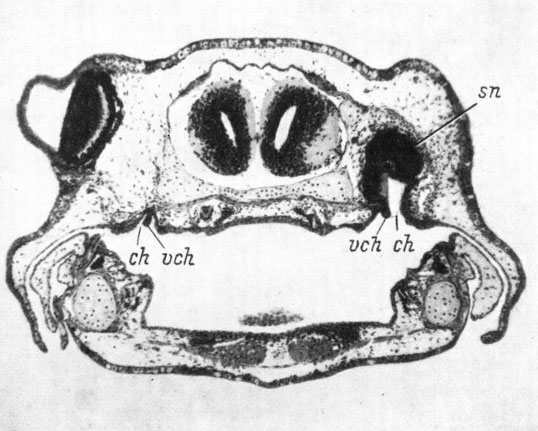
Рис. 40. Поперечный срез через область хоаны у оперированной личинки Hynobius keyserlingii длиной 31 мм. Слева орган обоняния отсутствует. (Увел. 38)
Для удобства сопоставления результатов операций у личинок различных исследовавшихся видов полученные данные сведены в табл. 3. Эта таблица наглядно показывает, что в отсутствие органа обоняния полностью нормальная хоана развивается только у представителей сем. Hynobiidae, в то время как высшие Urodela представляют группы с отсутствием хоаны, с сомнительными случаями и с образованием дефектной хоаны без клапана.
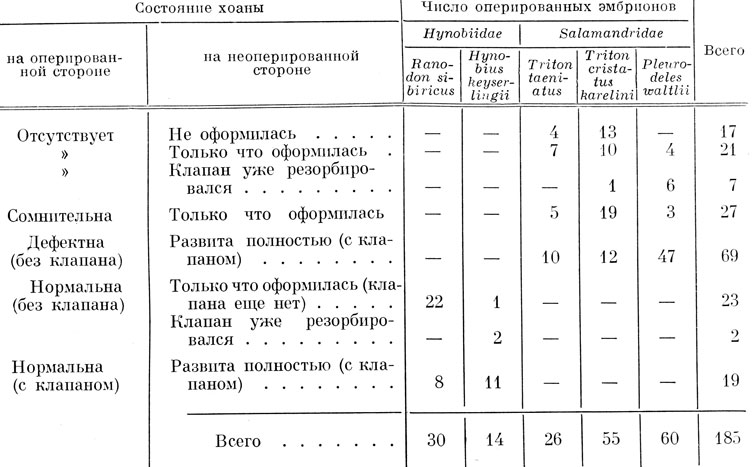
Таблица 3. Состояние хоаны на оперированной стороне в отсутствие обонятельного органа у хвостатых амфибий
Таким образом, полное удаление закладки органа обоняния у хвостатых амфибий не оказывает влияния на возникновение хоаны. Хоана, следовательно, образуется не за счет материала обонятельного мешка, а за счет эпителия стенки ротовой полости. Самостоятельное возникновение хоаны при отсутствии органа обоняния не означает, однако, что оно является полностью независимым: оно независимо только морфологически, так как выявился целый ряд данных, говорящих об индуктивном воздействии обонятельного мешка и его хоанального выроста на хоану. При полном отсутствии органа обоняния наблюдается следующее.
1. Более медленное развитие хоаны по сравнению с нормой у Salamandridae.
2. Более медленное образование хоанального клапана по сравнению с нормой у Hynobiidae.
3. Отсутствие хоанального клапана при вполне в остальном развитой хоане у Salamandridae.
4. Полное исчезновение хоаны во время метаморфоза у Salamandridae.
К перечисленным данным добавляются данные серии экспериментов с трансплантацией обонятельной плакоды об интенсивном индуктивном воздействии обонятельного мешка на эпителий глотки у Salamandridae и видимом отсутствии такого рода воздействия у Ranodoh sibiricus, у которого ни одного случая соединения хоанального выроста с выростом эпителия глотки не было (стр. 67).
При сопоставлении этих фактов получается чрезвычайно интересная картина, показывающая, что явно имеющаяся индуктивная связь между органом обоняния и хоаной гораздо слабее выражена у примитивных видов хвостатых амфибий. Действительно, у Hynobiidae в отличие от высших Urodela в отсутствие органа обоняния наблюдаются следующие отношения.
1. Хоана образуется почти одновременно с хоаной неоперированной стороны.
2. Развивается совершенно нормальная хоана с клапаном.
3. Во время метаморфоза хоана не исчезает.
Кроме того, пересадка обонятельной плакоды на место слухового пузырька у R. sibiricus ни разу не имела следствием возникновения ответной реакции эпителия глотки на индуктивное воздействие обонятельного мешка.
Перечисленные факты показывают, что у примитивных хвостатых амфибий наличие или отсутствие органа обоняния не имеет решающего значения для возникновения нормальной хоаны и сохранения ее во время наступления резорбционных процессов в метаморфозе, а следовательно, они не выявляют отчетливой формативной зависимости между этими образованиями. Единственным указанием на то, что такая зависимость (хотя и в очень малой степени) все же уже существует у самых примитивных Urodela является некоторое отставание в развитии хоанального клапана на оперированной стороне по сравнению с его развитием на неоперированной. Небольшая интенсивность индуктивного воздействия обонятельного мешка у Hynobiidae подтверждается и тем, что оно оказывается недостаточным для того, чтобы вызвать ответную реакцию чужеродного эпителия глотки при пересадке обонятельной плакоды на место удаленного слухового пузырька.
В то же время у Salamandridae в отсутствие органа обоняния отмечаются совсем иные закономерности.
1. Хоана образуется гораздо позже, чем на неоперированной стороне.
2. Никогда не образуется вполне нормальная хоана, так как клапан, ее закрывающий, всегда отсутствует.
3. Во время метаморфоза, когда в норме в области хоаны начинается процесс резорбции хоанального клапана, на оперированной стороне хоана оказывается втянутой в этот процесс и полностью исчезает.
Кроме того, пересаженная в область слухового пузырька обонятельная плакода способна вызвать ответную реакцию эпителия глотки в виде усиленного роста спиракулярного выроста по направлению к хоаналь-ному выросту обонятельного мешка.
Сравнение приведенных данных показывает, что у высших тритонов сем. Salamandridae для образования и полного развития хоаны необходимо присутствие, а следовательно, и индуктивное воздействие органа обоняния, в то время как у примитивных хвостатых амфибий сем. Hynobiidae оно требуется только в очень незначительной степени: отсутствие органа обоняния влечет за собой лишь некоторое отставание в развитии хоанального клапана по сравнению с его развитием на неоперированной стороне.
Таким образом, намечающаяся у низших хвостатых амфибий формообразовательная индуктивная связь между обонятельным мешком и хоаной заметно усиливается при переходе к высшим хвостатым амфибиям. Индуктивное формативное воздействие обонятельной плакоды у высших хвостатых амфибий возрастает настолько, что оказывается способным изменить направление разрастания спиракулярного выроста, глотки в сторону хоанального выроста обонятельного мешка.
Следовательно, в ряду хвостатых амфибий на примере органа обоняния с большой отчетливостью прослеживается возникновение и дальнейшее развитие морфогенетической коррелятивной связи типа эмбриональной индукции, причем эта индуктивная связь является еще в значительной степени несовершенной даже у высших представителей отряда: в случае полного отсутствия индукционного воздействия хоана все-таки образуется, хотя и без клапана, а наличие частичного регенерата обонятельного мешка, не соединенного с ротовой полостью, не оказывает заметного влияния на образование хоаны (при частичной регенерации обонятельного мешка результаты получаются такие же, как и при полном отсутствии органа обоняния, за исключением очень небольшого ускорения развития хоаны, наблюдающегося при наличии регенерата мешка).
Из сказанного следует, что в пределах отряда хвостатых амфибий формообразовательная система (обонятельная закладка - хоана) находится еще только в процессе становления. Этот сам по себе интереснейший факт, относящийся к проблеме исторического возникновения формообразовательных механизмов, заслуживающей отдельного рассмотрения, имеет для исследуемого вопроса чисто прикладное значение: он говорит о том, что упомянутая система образовалась сравнительно недавно (не в смысле абсолютной протяженности геологического времени, а в смысле интенсивности филогенетических процессов за это время). Это дает нам право предполагать, что между обонятельной закладкой и хоаной ранее существовали иные отношения, послужившие базой для возникновения формообразовательной индуктивной связи (Медведева, 1963а).
Таким образом, в условиях эксперимента у хвостатых амфибий хоана возникает морфологически независимо при полном отсутствии органа обоняния, причем она развивается не из ткани обонятельного мешка, который в этих условиях отсутствует, а из эпителия стенки ротовой полости. Однако нельзя сказать, что образование хоаны хвостатых амфибий совершенно независимо от органа обоняния, так как хотя наличие последнего не является непременным условием возникновения хоаны, все же между ними существует формообразовательная связь типа эмбриональной индукции, еще очень слабо выраженная у представителей примитивного семейства Hynobiidae и значительно усиливающаяся у высших тритонов.
Сравнивая эти данные с результатами экспериментальных исследований процесса образования хоан у бесхвостых амфибий, можно заключить, что как у Anura, так и у Urodela наблюдается морфологически независимое от обонятельного мешка развитие хоаны. Но поскольку хоана является у наземных позвоночных неотъемлемой частью органа обоняния в целом, это означает, что орган обоняния хвостатых и бесхвостых амфибий имеет совершенно особый, не такой, как у других Tetrapoda, способ развития из двух морфологически обособленных источников, причем в некоторых случаях эти источники даже могут относиться к разным зародышевым листкам.
Установленная у хвостатых амфибий формообразовательная индуктивная связь между двумя морфологически обособленными закладками органа обоняния, очевидно, существует и у бесхвостых амфибий: об этом говорит образование у них в отсутствие обонятельного мешка в большинстве случаев не полноценной, а дефектной хоаны с недоразвивающимися клапанами. Изменений в степени индуктивного воздействия обонятельного мешка на хоану, отчетливо выявившихся в ряду хвостатых амфибий, у бесхвостых амфибий проследить не удалось; они показали в этом отношении одинаковые у всех изучавшихся видов результаты, не сходные с результатами, полученными у хвостатых амфибий: образование, как правило, дефектной хоаны, а в виде исключения - нормальной. Следовательно, по состоянию формообразовательной системы обонятельная закладка - хоана исследованные бесхвостые амфибии находятся на промежуточной ступени между теми состояниями этой системы, которые наблюдаются у примитивных и высших хвостатых амфибий, поскольку у всех Anura, как более примитивных, так и высоко организованных, отсутствие индуктивного влияния органа обоняния оказывает отрицательное действие на развитие хоаны, но еще не всегда - в отдельных случаях хоана сохраняет способность к нормальному независимому формообразованию. Однако эти различные состояния формообразовательной системы обонятельная закладка - хоана у бесхвостых и хвостатых амфибий исходят из общей для тех и других основы - предшествовавшего независимого от органа обоняния развития хоаны.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'