А. Трансплантация обонятельной плакоды на место удаленного слухового пузырька
Изложенное выше описание развития хоанального канала (глава II) подтверждает данные Гинсберга (Hinsberg, 1901) и других исследователей об образовании его у хвостатых и бесхвостых амфибий не за счет оро-назальной борозды, а за счет выроста самого обонятельного мешка. Однако окончательно этот спорный вопрос может быть решен только при помощи эксперимента. Одним из возможных путей является пересадка обонятельной плакоды в другое место, так как в этом случае образование хоанального канала подтвердит его возникновение из материала самой плакоды, отсутствие же хоанального канала должно обнаружить соответствующее отсутствие материала для его образования.
С этой целью была поставлена серия опытов по гомотрансплантации обонятельной плакоды на место удалявшегося слухового пузырька, которое удобно тем, что в случае образования хоанального канала последний имеет возможность соединения с располагающейся вентральнее глоткой.
Бесхвостые амфибии. Подопытным материалом послужили эмбрионы зеленой и травяной лягушек, чесночницы и зеленой жабы на стадиях ранней, средней и поздней хвостовой почки, когда орган обоняния представляет собой обонятельную плакоду или очень мелкую обонятельную ямку. Доноры и реципиенты брались одного возраста. Видовая принадлежность не оказала влияния на конечные результаты. Всего было прооперировано 102 эмбриона, из них выжило 91, трансплантаты прижились у 83 экз. Большая часть материала зафиксирована на стадии образования жаберной складки, когда в норме у личинок уже имеются хорошо развитые хоаны. Остальные личинки фиксировались на промежуточных стадиях с целью выяснения хода развития. Окраска производилась тотально борным кармином.
Изучение серий поперечных срезов оперированных личинок дало следующие результаты.
1. В 8 случаях обонятельный орган оказался недостаточно развившимся (фиксация на промежуточных стадиях).
2. В 22 случаях обонятельный мешок образовал выросты хоанального типа. В 18 из этих случаев наступило соединение этих выростов с глоткой или же с жаберной полостью.
3. В остальных 53 случаях образования хоанального выроста не произошло.
Для выяснения причин отсутствия образования хоанального выроста в последней группе случаев было проведено сравнение обонятельных органов, развившихся из пересаженных плакод, с восковыми моделями и графическими реконструкциями их на той же стадии нормального развития и установлено, что в 30 случаях трансплантаты были ориентированы неправильно, в результате чего орган обоняния оказался перевернутым на 90°, 180 или 270° (при переносе трансплантата очень трудно сохранить его правильную ориентировку). В остальных 23 случаях имело место неправильное дефектное развитие органа обоняния, сделавшее невозможным установление его ориентировки. Следовательно, во всех случаях отсутствия хоанального выроста орган обоняния либо неправильно развивался, либо был неправильно ориентирован. Случаев правильной ориентировки органа в этой группе не было.
В 22 случаях второй группы, т. е. в 26.5% от общего числа личинок с прижившимися трансплантатами, наблюдалось образование выростов хоанального типа из ткани самого обонятельного мешка, причем эта группа включает личинок всех четырех исследованных видов. В 4 случаях имело место образование обонятельным мешком нескольких выростов. В 18 случаях этой группы установилось соединение выроста со стенкой кишечной трубки, причем в 8 из них непосредственно со стенкой глотки, имеющей в этом участке определенно энтодермальное происхождение. В ряде случаев наблюдалось возникновение обонятельного нерва и установление связи с мозгом и со слуховым ганглием, однако это наблюдалось и в третьей группе случаев, в которой не было образования хоанальных выростов. Образования канала, т. е. полости в выросте, и собственно хоаны не было ни в одном случае. В 11 случаях пересаженные обонятельные органы имели правильную ориентировку, в остальных случаях ее правильность не удалось установить из-за отсутствия латерального аппендикса и якобсонова органа. На рис. 35 даются графические реконструкции обонятельного органа, развившегося на месте слухового пузырька.
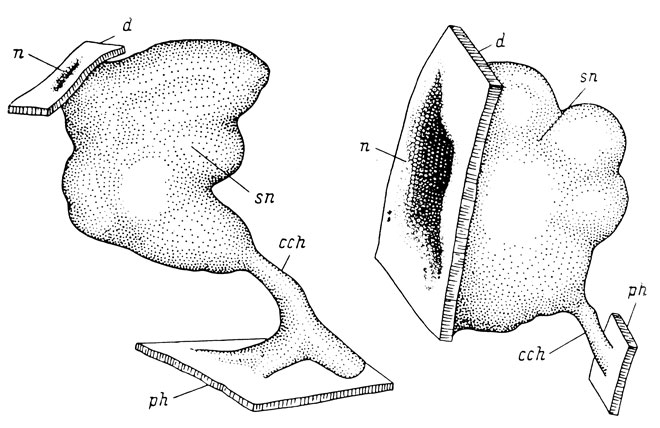
Рис. 35. Графические реконструкции органа обоняния Rana esculenta, пересаженного на место слухового пузырька
Результаты эксперимента наглядно показали, что материалом для образования хоанального канала служат клетки самого обонятельного мешка. Если бы хоанальный канал был обязан своим возникновением оро-назальной борозде, превращающейся в эпителиальный тяж, погружающийся под эктодерму, как утверждала М. Курепина (1926, 1931), образование хоанальных выростов в условиях данного эксперимента не могло бы иметь места. Случаи отсутствия хоанального выроста вполне объяснимы неправильным развитием органа в чуждом месте или его неправильной ориентировкой. Соединение хоанального выроста в ряде случаев непосредственно с энтодермальной стенкой глотки хорошо согласуется с данными по нормальному развитию хоанального канала бесхвостых амфибий, изложенными выше (см. главу II).
Хвостатые амфибии. Трансплантация обонятельной плакоды на место удаленного слухового пузырька была проведена у эмбрионов обыкновенного, кавказского подвида гребенчатого и испанского тритонов, а также у эмбрионов Ranodon sibiricus. Операции производились у обыкновенного тритона на стадиях 24 - 29 по Глезнеру, у кавказского подвида гребенчатого тритона и R. sibiricus - на стадиях, соответствующих указанным у обыкновенного тритона, у испанского тритона - на стадиях 24 - 29 по Гальену и Дюроше. У первых трех видов доноры и реципиенты брались одинакового возраста, у испанского тритона пересадки производились по способу аутотрансплантации. Личинки выращивались до стадий несколько более поздних, чем та, на которой в норме образуются хоаны, т. е. до стадий 40, 41 по Глезнеру у обыкновенного тритона и соответственно этим стадиям у остальных видов. Объекты фиксировались, окрашивались тотально борным кармином и гемакальцием, а на стеклах квасцовым гематоксилином и по Маллори.
Из 150 выращенных, зафиксированных и исследованных личинок трансплантаты прижились у 109 экз. Поскольку конечные результаты в отличие от таковых у бесхвостых амфибий оказались зависимыми от систематического положения, следует привести распределение этого количества по отдельным видам: обыкновенный тритон - 8, кавказский подвид гребенчатого тритона - 45, испанский тритон - 29, R. sibiricus - 27 личинок. Изучение серий поперечных срезов оперированных личинок дало следующие результаты.
1. В 13 случаях обонятельная плакода не развивалась и дегенерировала.
2. В 39 случаях развившийся из обонятельной плакоды обонятельный мешок образовал выросты, которые в 17 случаях ничем не отличались от хоанальных выростов, возникающих при нормальном развитии. В 9 из этих 17 случаев осуществилось или почти осуществилось соединение выростов с расположенной вентральнее места пересадки глоткой.
3. В остальных 57 случаях обонятельная плакода превратилась в обонятельный мешок, но никаких выростов при этом не образовывалось.
Как и у бесхвостых амфибий, в двух последних группах имеются случаи образования обонятельного нерва и последующего соединения его либо с мозгом, либо со слуховым ганглием (соединение трансплантированной обонятельной закладки с мозгом описано многими авторами: Bell, 1907; Burr, 1924 - 1925; Teichmann, 1955, и др.). Следовательно, возникновение обонятельного нерва и соединение его с мозгом не оказывают влияния на образование хоанального выроста, этот процесс детерминирован ранее.
Вторая группа объединяет случаи, давшие положительные результаты, но из них 9 случаев с осуществлением соединения хоанального выроста с глоткой заслуживают выделения в отдельную подгруппу, поскольку они относятся не ко всем исследованным видам, а только к трем представителям сем. Salamandridae. у R. sibiricus - представителя примитивного сем. Hynobiidae хоанальный вырост обонятельного мешка образовывался, но соединения этого выроста с глоткой не наблюдалось ни разу. Общее число положительных случаев достаточно высоко и показывает, что хоанальный вырост образуется из ткани самого обонятельного мешка, а не из оро-назальной борозды, так как при пересадке обонятельной плакоды особое внимание обращалось на то, чтобы избежать захвата покровного эпителия, лежащего между плакодой и местом будущего ротового впячивания. В некоторых случаях возникли выросты, настолько похожие на образующийся в норме хоанальный вырост обонятельного мешка, что в природе их не остается никаких сомнений (рис. 36), причем образование таких выростов имело место даже в тех случаях, когда обонятельный мешок оказывался повернутым на 180°.
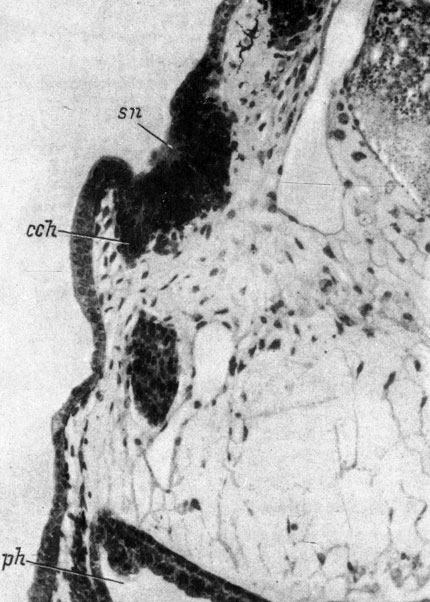
Рис. 36. Поперечный срез через обонятельный мешок, пересаженный в область слухового пузырька, у личинки Ranodon sibiricus длиной 16.5 мм. (Увел. 100)
Сравнительно небольшое число случаев, когда наступило соединение хоанального выроста обонятельного мешка с глоткой, по сравнению с результатами таких же пересадок у бесхвостых амфибий, можно объяснить тем, что в норме обонятельный мешок хвостатых амфибий образует очень короткий хоанальный вырост, так как каудальная половина хоанального канала развивается за счет хоанального выроста передней кишки (см. главу II). Поэтому осуществление соединения с глоткой, расположенной на довольно большом расстоянии от пересаженной на поверхность тела обонятельной плакоды, в данном случае более затруднено, чем в случае образования хоанального выроста обонятельного мешка при трансплантации плакоды у бесхвостых амфибий.
Чрезвычайно интересно, что в тех случаях, когда происходит соединение хоанального выроста обонятельного мешка с глоткой, оно становится возможным благодаря тому, что сама глотка образует вырост, направленный навстречу хоанальному выросту обонятельного мешка (рис. 37). Этот вырост глотки имеется и в норме, но не во всех случаях, и у разных видов достигает различной степени развития. Судя по его положению под передней частью развивающегося внутреннего уха и у каудального края нёбно-квадратного хряща, он представляет собой остаток спиракулярной щели. Однако, во-первых, в норме этот вырост глотки никогда не достигает такой степени развития, как в условиях данного эксперимента, а во-вторых, в последнем случае он растет точно в направлении образующегося хоанального выроста обонятельного мешка, а не в сторону слухового пузырька, как обычно. Следовательно, здесь с несомненностью выявляется индуктивное воздействие со стороны обонятельного мешка и его хоанального выроста и ответная реакция эпителия глотки. Следует еще раз подчеркнуть, что у R. sibiricus не было ни одного случая соединения хоанального выроста обонятельного мешка с глоткой и соответственно роста выроста глотки в направлении трансплантированного обонятельного мешка. Эти данные, по-видимому, указывают на большую индукционную способность обонятельного мешка высших хвостатых амфибий по сравнению с Hynobiidae.
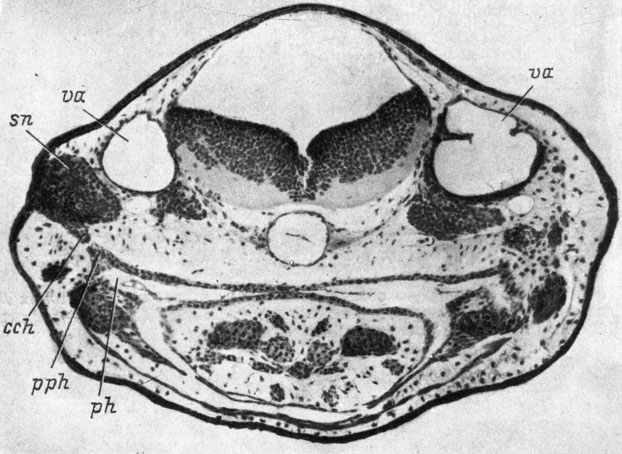
Рис. 37. Поперечный срез головы личинки Triton cristatus karelini длиной 10.5 мм с обонятельным мешком, пересаженным в область слухового пузырька. (Увел. 76)
Подводя итоги опытов по пересадке обонятельной плакоды в область слухового пузырька у хвостатых амфибий, можно заключить, что у них так же, как у бесхвостых, хоанальный канал, а именно его ростральная половина, возникает не из оро-назальной борозды, а за счет выроста ткани самого обонятельного мешка.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'