Колебания численности и их причины
Изучение и вскрытие причин, определяющих колебания численности животных в природе, является одной из важнейших задач современной зоологии. Эти изменения определяют динамику биомассы вида, а следовательно, и степени участия вида в работе биогеоценозов. Первично количественные изменения числа особей в популяциях представляют собой итог воздействия, факторов биотического и абиотического характера. При этом надо различать истинные изменения численности и временные, которые могут быть вызваны миграциями животных из данных, биотопов в связи с неблагоприятными условиями (засуха, наводнение) или различием периодов активности разных возрастных и половых групп. Наблюдения показывают, что как биотические, так и абиотические факторы оказывают особенно массированное воздействие на численность наземных позвоночных, если они связаны с антропогенным воздействием.
Периодические колебания численности
Периодические колебания численности населения ящериц в основном связаны с размножением и возрастной смертностью, т. е. являются сезонными. В ряде случаев количественные флуктуации численности в популяциях могут быть вызваны периодичностью вспышек кормовых видов и периодических изменений "давления" хищников.
Известно, что всякая популяция имеет свою специфическую ритмику численности (как по полу, так и по возрасту). К сожалению, имеются лишь отрывочные данные по ритмике численности особей разного пола прыткой ящерицы по сезонам. В. К. Жаркова (1973а) исходя из факта, что количественные соотношения разных возрастных групп в популяции иные, на основе имеющегося у нее материала определяет различие в возрастной смертности самцов и самок (табл. 57).
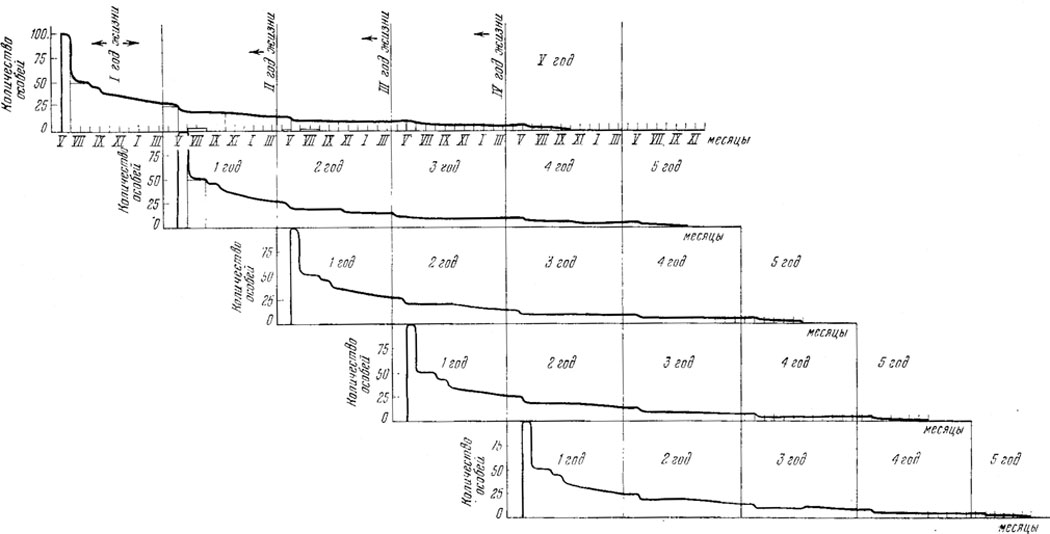
Рис. 92. Схема движения численности 5 последовательных поколений модельной популяции прыткой ящерицы. Исходная численность каждого поколения - 100 особей (отложенных яиц). По данным А. С. Баранова, А. В. Яблокова, А. В. Валецкого (см. также рис. 87)
Как видно из табл. 57, за первый и второй годы жизни среди ящериц Мещерской низменности самцов погибает почти в два раза больше, чем самок. Лишь на третьем году жизни резко увеличивается процент гибели самок. Такой ход динамики численности приводит к преобладанию в популяции молодых самок. В более старших возрастных группах, наоборот, происходит интенсивная смертность самой, и соотношение полов выравнивается. Возможно, именно такая дифференцированная смертность объясняет несколько большую численность самок в некоторых природных популяциях.
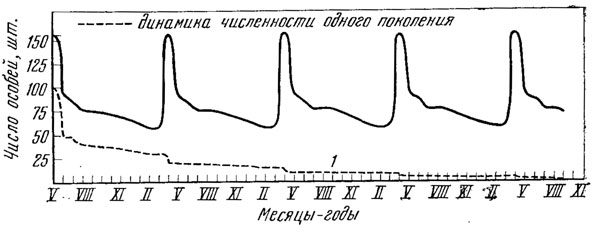
Рис. 93. Обобщенная схема динамики численности модельной популяции прыткой ящерицы. Исходная численность - 156 особей - определена путем суммирования разных возрастных групп трех поколений (см. рис. 87)
Ритмика колебаний численности особей разного пола подчиняется тем же общим закономерностям, которые действуют при возрастных колебаниях, с некоторыми поправками для разных полов. В настоящее время можно построить модель динамики численности населения прыткой ящерицы, определяемую возрастной смертностью (рис. 92). Как показано в гл. XV, темп смертности животных на протяжении их жизни оказывается различным. Проследим динамику численности одного поколения. Принимая данные табл. 62 (гл. XV) за исходные, получаем, что наибольшая численность в популяции достигается в момент откладки яиц (отложенное оплодотворенное яйцо - это уже особь) и выхода молодых. Учитывая большую смертность среди новорожденных (50%), которые гибнут до зимовки и после нее, к следующей весне в живых останется всего 25% от родившихся этого поколения. На второй год погибнет около 33% от достигших возраста в один год. На третий год жизни смертность, видимо, несколько снижается, но на четвертом и пятом годах жизни достигает соответственно 50 и 100% от числа доживших до этого возраста особей.
Таблица 57
| Генерация | Самцы (исходная численность сеголетков 1-й генерации 272 экз.) | Самки (исходная численность сеголетков 1-й генерации 203 экз.) | ||
| число особей | % | число особей | % | |
| 1-я | 82 | 30,0 | 34 | 16,7 |
| 2-я | 118 | 67,3 | 45 | 26,6 |
| 3-я | 49 | 68,0 | 73 | 58,8 |
*(Под генерацией в данном случае имеется в виду потомство, появившееся на свет в один сезон размножения.)
Суммарная картина сезонных колебаний численности в популяции прыткой ящерицы будет намного сложнее, так как в каждый момент времени в популяции будут находиться особи пяти поколений, каждое из которых имеет свой темп смертности. В результате модель численности популяции прыткой ящерицы будет выглядеть следующим образом (рис. 93). Необходимо подчеркнуть, что в этой модели учтены только сезонная циклика; на нее в природе практически всегда накладывается апериодическая цикличность.
Таблица 58
| Биотоп | 1965 г. | 1966 г. | 1967 г. | 1968 г. |
| Смешанный лес | 4,6 | 5,0 | 4,8 | 5,2 |
| Сосновые посадки | - | - | 5,5 | 4,1 |
| Разнотравный луг | 1,3 | 1,1 | 1,1 | 1,6 |
| Суходольный луг | - | 1,3 | 1,0 | 0,6 |
| Берег реки | 4,3 | 4,6 | 4,7 | 4,8 |
Безусловно, что в разных частях ареала динамика изменений сезонной численности популяции должна быть различной. Многолетняя динамика численности представляет собой итог сезонных динамик. Но это не простая сумма, а как бы наложение друг на друга различных часто противоположно направленных явлений, обусловленных различными факторами. Говоря о сочетании факторов, имеются в виду как биотические, так и абиотические факторы, действующие на природные популяции. Проиллюстрируем колебания численности в разные годы на примере пяти групп ящериц в северной лесостепи Европейской части СССР (табл. 58).
Как видно, каждому биотопу присуща определенная динамика колебаний численности. Так, на склонах реки численность ящериц за эти четыре года постоянно увеличивается, в то (время как в сосновых посадках ящерица отсутствует, затем резкий "всплеск" численности с последующим сокращением численности населения. В смешанном лесу, на разнотравном лугу происходят изменения численности другого порядка.
Несомненно, колебания численности в популяциях прыткой ящерицы, связанные с действием биотических факторов, могут быть вызваны либо периодическими вспышками численности основных кормов (см. гл. VI), либо периодичностью давления пресса хищников, либо, наконец, резкими вспышками численности конкурирующих видов.
Увеличение численности кормовых видов, безусловно, приводит к некоторому увеличению численности популяции (при условии отсутствия пресса хищников и конкурирующих видов), падение же численности пищевых видов в некоторых случаях приводит к тому, что взрослые особи начинают пожирать молодых (каннибализм), тем самым сокращая численность популяции. В некоторых случаях происходит ложное сокращение численности популяции в результате миграции. Такие миграции могут быть вызваны увеличением численности популяций конкурирующих видов или резким спадом численности кормовых видов. Иногда популяции мигрируют полностью и переходят в биотопы, совершенно ей не свойственные, в стадии переживания (см. гл. IX). Важно добавить, что кормовые виды и виды-конкуренты, видимо, не имеют столь же существенного значения в регуляции численного состава популяций прыткой ящерицы (Лукина, 1966; Тертышников, 1972а, б; и др.)
Немаловажную роль в колебаниях численности популяций играют хищники. Резкое увеличение или падение численности хищников неизбежно приводит к сокращениям или увеличениям численности популяций прыткой ящерицы. В то же время значение хищников как регуляторов численности популяций прыткой ящерицы крайне недостаточно изучено. По расчетам М. Ф. Тертышникова, в Ставропольском крае, на стационарном участке, расположенном в верховьях р. Томузловки, за сезон гибнет 37,2% общей биомассы популяции ящерицы в результате воздействия на них учтенных фоновых врагов из позвоночных. В этом районе пресс хищников является дополнительной причиной, задерживающей рост численности в популяции. "Давление" хищников, несомненно, различно и в разных популяциях. Об этом косвенно свидетельствуют данные в гл. XIII о различной доле особей с регенерированными хвостами (т. е. особей, подвергавшихся нападению хищников и удачно их избежавших). Напомним лишь, что в некоторых популяциях больше половины взрослых особей несут на себе следы таких нападений.
В целом можно сказать, что периодические колебания, видимо, не вызывают изменений численности популяций прыткой ящерицы более чем на один порядок.
Апериодические колебания численности
Факторы, способствующие массовому размножению ящериц популяции, и факторы, определяющие наступление следующего за ним периода депрессии численности, могут быть вызваны апериодическими явлениями. Такими явлениями могут быть катастрофические изменения биогеоценозов в результате пожаров, наводнений, засухи, сильных морозов в малоснежные зимы и прочие явления, прямо или косвенно благоприятствующие или, наоборот, препятствующие росту численности. Колебания численности могут быть связаны и с длительными изменениями природных условий, вызванными деятельностью человека (опустынивание, засоление больших территорий и т. п., или длительными изменениями климата типа ледниковых периодов. Иногда такого рода причины могут приводить к массовой гибели животных, тем более что прыткая ящерица, являясь пойкилотермным животным, весьма зависит от климатических условий.
По мнению многих авторов (Терентьев, 1946; Лукина, 1966б; Гаранин, 1971, Тертышников, 1972б; Жаркова, 1973а; и др.), основное сокращение численности ящериц происходит в основном за счет гибели яиц. Если принять все отложенные яйца в популяции за 100%, то к моменту выхода молодых гибнет от 40 до 60%. Этот процент резко увеличивается особенно в дождливое и холодное лето (Гаранин, 1971). Другой критический момент, связанный с физическими факторами среды, в жизни прыткой ящерицы - осень с ранними заморозками. Именно в этот период времени, когда животные собираются уходить на зимнюю спячку, такого рода явления наиболее опасны (особенно для молодых этого года рождения, которые позднее уходят на зимовку).
Наконец, последним критическим моментом, влияющим на численность популяций, является зимовка. Численность особенно резко падает в морозные малоснежные зимы (Гаранин, 1971; Тертышников, 1972б). Обычно взрослые ящерицы зимуют в своих норах или в старых норах грызунов (см. гл. V). Если в открытых биотопах норы большинства грызунов глубокие и как зимние убежища являются надежными (так как температура в них не опускается ниже 0°), то этого нельзя обычно сказать о норах, вырытых самими ящерицами, обычно неглубоких, а также о трещинах и щелях, где часто в почве зимуют молодые особи. В малоснежные холодные зимы, когда почва недостаточно покрыта снегом, а следовательно нарушается температурный режим в зимних норах, многие животные, зимующие в таких норах, погибают. Поэтому значительно более надежными для выживаемости ящериц зимой являются лесные биотопы, лесополосы, придорожные канавы, овраги и т. п., так как именно в этих местах скапливается значительное количество снега, утепляющего почву. Здесь массовая гибель животных в морозные зимы может наступить лишь в исключительных случаях. В. И. Гаранин (1971) приводит такой пример. Малоснежная суровая зима 1968 г. в Волжко-Камском заповеднике привела к резкому сокращению численности ящериц. Именно во время зимовки происходит резкое сокращение численности среди молодых животных в популяции. Так, М. Ф. Тертышников показал, что отход яиц и смертность сеголетков в первую зимовку в Ставропольской возвышенности составляет 25,7%.
Наиболее существенные изменения среды обитания, носящие катастрофический характер, приводят к резкому снижению ее численности. К типичным явлениям такого порядка можно отнести пожары, наводнения и засухи. По сведениям В. К. Жарковой (1973а), в 1967 г. после сильных дождей в районе Окского заповедника реки вышли из берегов и затопили биотопы, занимаемые прыткой ящерицей. Так, в пойменном лугу и в пойменных посадках сосны на р. Выше в этот год наблюдалось резкое сокращение численности прыткой ящерицы: на пойменном лугу численность ящериц в среднем сократилась в 4 раза по сравнению с 1966 г., а в пойменных посадках в 2,5 раза. Надо заметить, что при этом могут совершенно исчезнуть отдельные демы или целые их группы. Но популяции в большинстве случаев сохраняются, хотя и могут при этом достигать минимальных численностей, достаточных все же для дальнейшего существования.
Наиболее существенные изменения среды, связанные с длительным изменением природных условий, в настоящее время чаще бывают связаны с антропогенными воздействиями. Например, одной из причин гибели прытких ящериц является создание новых водохранилищ на пойменных землях. На берегах Куйбышевского водохранилища Гаранин (1971) отмечает резкое сокращение численности рептилий и в том числе прыткой ящерицы. В то же время на некоторых островах водохранилища сократившиеся сначала численно популяции затем заметно увеличились.
Более серьезным и распространенным фактором, отрицательно влияющим на численность ящериц, является неумеренное использование ядохимикатов в сельском и лесном хозяйстве. Существует строгая корреляция между обработкой ядохимикатами участков Мещеры и населенностью их ящерицами (Жаркова, 1973б). В этом районе не заселенные прыткой ящерицей, но подходящие для них биотопы составляют от 32 до 49% обследованной территории. Есть все основания согласиться с предостережением Е. Рене (Rene, 1969) и К. Корбетта (Corbett, 1969) об опасности вымирания прыткой ящерицы в развитых промышленных районах в связи с полным антропогенным разрушением естественных биотопов.
Есть и другая сторона антропогенного влияния на численность прыткой ящерицы. Наблюдения, проведенные в 1970 - 1974 гг. в разных частях ареала прыткой ящерицы, показывают, что она в некоторых районах становится "антропогенным видом". Широкая способность этого вида приспосабливаться к антропогенным биотопам (см. табл. 5) несомненно позволит этому виду не только "не сокращать свою численность при контактах с цивилизацией, но, возможно, в некоторых частях своего ареала даже увеличивать ее. Пока же в результате активного антропогенного воздействия численность прытких ящериц резко сокращается на Ставропольской возвышенности (Тертышников, 1972в), в Калужской обл. (Стрельцов), во многих районах Сибири (Баранов и др., сообщ.), Латвийской (Бахарев, 1971) и Эстонской ССР (Вельдре, личное сообщ.), а в окрестностях г. Махачкала (Хонякина, Кутузова, личн. сообщ.) прыткая ящерица исчезла вообще.
Таким образом, в настоящее время лишь при очень резких изменениях среды, обычно, связанных с разрушением биоценозов под воздействием человека, численность популяций сокращается ниже критического уровня, за которым следует вымирание популяции.
Периодические (сезонные и годовые) колебания численности прыткой ящерицы, видимо, не превышают одного порядка величин, тогда как апериодические колебания оказываются часто более значительными.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'