Определение возраста
При исследовании большой единовременно взятой выборки можно определить относительный возраст особи по ее положению в ряду размерного состава. Однако эта методика дает тем более надежный результат, чем более сжаты сроки размножения. Учитывая, что у прыткой ящерицы сроки размножения достаточно растянуты, а также и то, что индивидуальная изменчивость темпа роста может быть велика, следует с осторожностью интерпретировать выделяемые группы как годовые классы. Ниже будет показано, что четко выделяется только первая возрастная группа (сеголетки - летом и осенью и перезимовавшие один раз - весной). Особей старших возрастных групп нельзя наверняка отнести к определенной возрастной группе. Поэтому проблема разработки методов точного определения абсолютного возраста ящериц становится актуальной.
Четкая методика определения абсолютного возраста рептилий до сих пор отсутствует. Обычно применяемое разделение на половозрелых и неполовозрелых по состоянию половых желез по сути не является определением возраста. Выделение возрастных классов по размерам тела (Залежский, 1938; Банников, 1940; Щепотьев, 1948; Терентьев, Чернов, 1949; Банников, Денисова, 1956; Богданов, 1965; Blanchard, Blanchard, 1931; Hamilton, 1934; Michael, 1972; Cruce, 1971; Peters, 1964; Gibbons, 1972; и др.) нельзя считать удовлетворительным прежде всего потому, что по достижении половой зрелости темп роста животных сильно снижается и чем дальше, тем медленнее увеличиваются размеры тела. Это ведет к тому, что в последнюю размерную группу из тех, которые можно выделить по длине тела, входят взрослые животные разного возраста. Кроме того эта методика не учитывает индивидуальных различий скорости роста. Было предпринято довольно много попыток определения возраста рептилий по слоям в кости. То, что в костях рептилий, так же как и остальных позвоночных, имеются слои, отражающие периодичность роста животных, известно давно. Слоистая структура кости видна на тонких плоских костях или на тонких шлифах костей (просветленных в глицерине или других просветляющих жидкостях), а также на срезах кости, окрашенных гематоксилином. Узкие линии, образующиеся во время задержки роста и разграничивающие широкие слои костной ткани, называют линиями задержки роста, или линиями склеивания и по их числу определяют возраст животного (Клевезаль, Клейненберг, 1967). Еще в 1907 г. была описана слоистость периостальной кости в диафизе у нескольких видов вымерших и современных рептилий (Seitz, 1907). Тогда же было высказано предположение, что широкие зоны костной ткани отлагаются летом, а узкие линии в холодное время года. Затем слои в кости рептилий были обнаружены целым рядом исследователей у разных видов рептилий (Wallis, 1928; Емельянов, 1929; Mattox, 1935; Брюзгин, 1939; Peter-Rousseaux, 1953; Saint-Girons, 1957, 1965; Peabody, 1958, 1961; Thireau, 1967; Castanet, 1974; и др.).
Сопоставление размеров тела животного с числом слоев в кости дало исследователям основание полагать, что число слоев в кости соответствует их возрасту. Однако никто из исследователей не приводил достаточно веских доказательств того, что видимые слои действительно годовые.
Некоторые авторы выступили с критикой методики определения возраста рептилий по слоям в кости (Dobie, 1971; Griffiths, 1961). Дж. З. Доби (Dobie, 1971), сравнивая картину слоистости на щитках карапакса черепах (подсчет числа слоев на щитках карапакса - общепринятая методика определения возраста черепах) со слоистостью, видимой на срезах ее позвонков и нижней челюсти, обнаружил лишь очень слабую положительную корреляцию числа слоев на щитках и срезах костей. Гриффите (Griffiths, 1961), изучавший слои в костях змей, возраст которых был известен, так как их инкубировали и выращивали в неволе в условиях, близких к естественным, сделал вывод о ненадежности этой методики. Одним из его возражений было то, что в длинных костях кроме отложения новых слоев костной ткани со стороны периоста, в процессе роста со стороны эндоста идет разорбция ранее отложившейся кости, и первые слои не сохраняются.
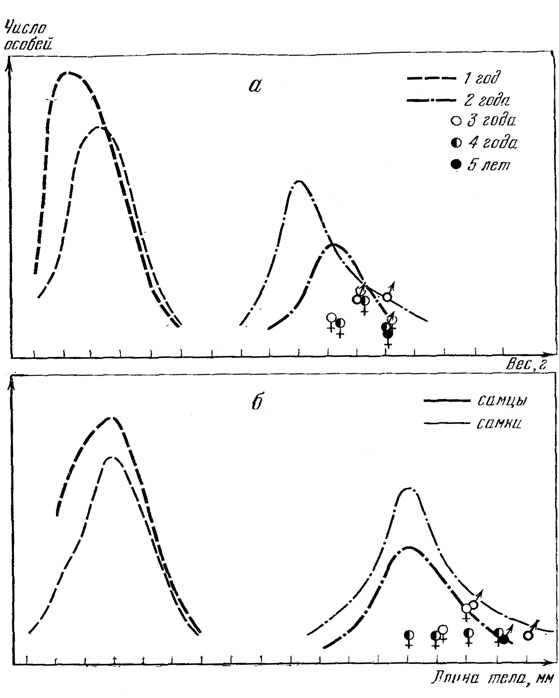
Рис. 76. Распределение особей по весу (а) и длине тела (б) в соответствии с возрастом, определенным по числу слоев в костях. Выборка из популяции окр. г. Умани, УССР 19 мая 1970 г.
Для решения вопроса о возможности определения возраста рептилий по слоям в кости идеальным было бы провести исследование животных точно известного возраста, взятых из естественных условий. Второй путь - исследование большой, единовременно взятой из естественных условий выборки. Поскольку у нас не было животных известного возраста, мы выбрали второй путь.
В нашем распоряжении была выборка прыткой ящерицы (93 экз.), добытых в середине мая 1971 г. в районе г. Умани, и 33 экз. сеголетков, добытых в августе того же года в Новгородской области.
Для изучения закономерности образования слоев в костях прытких ящериц были изготовлены поперечные срезы из середины диафизов плечевой и бедренной костей. Кости декальцинировали и срезы окрашивали гематоксилином Эрлиха, пользуясь методикой, которая была применена при определении возраста амфибий (Клейненберг, Смирина, 1969).
В уманской выборке по длине тела нами были выделены только 2 группы: годовики и взрослые ящерицы (рис. 77.). Первая возрастная группа, которую можно выделить и по длине тела и по картине на поперечных срезах кости - это сеголетки. Длина тела сеголетков, имевшихся в нашем распоряжении, колебалась от 29 до 42 мм. Картина, видимая на срезах, была неодинаковой. У особей длиной тела от 29 до 32 мм в кости не было видно ни одной линии склеивания, а у особей длиной тела от 32 до 42 мм была видна линия склеивания.
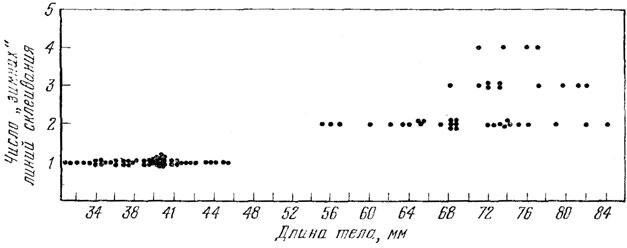
Рис. 77. Соотношение длины тела и числа 'зимних' линий склеивания в кости прыткой ящерицы Допуляция окр. г. Умани, УССР
У особей второй возрастной группы, которую мы выделяем из уманьской выборки, длиной тела от 31 до 47 мм, видны две линии склеивания (рис. 78, I). Этих ящериц можно считать зимовавшими один раз, или годовиками.
Очевидно, у сеголетков в первое лето жизни рост идет неравномерно; как правило, у них бывает одна задержка роста, которая и характеризуется первой линией склеивания. Вторая линия образуется в результате зимней задержки роста.
Следующая возрастная группа, которую можно выделить по картине на поперечных срезах костей, это ящерицы с двумя группами линий склеивания (см. рис. 78, II). У них около костномозговой полости обычно хорошо видны две частично резорбированные линии склеивания, сближенные одна с другой, которые мы считаем первой группой линий. Это, очевидно, следы от линии, которая имелась еще у сеголетков и от первой зимовки. Далее виден большой прирост кости и ближе к наружному краю еще одна, часто двойная или даже тройная линия склеивания (вторая группа линий). Ящериц с двумя группами линий склеивания можно считать зимовавшими два раза, т. е. двухгодовиками.
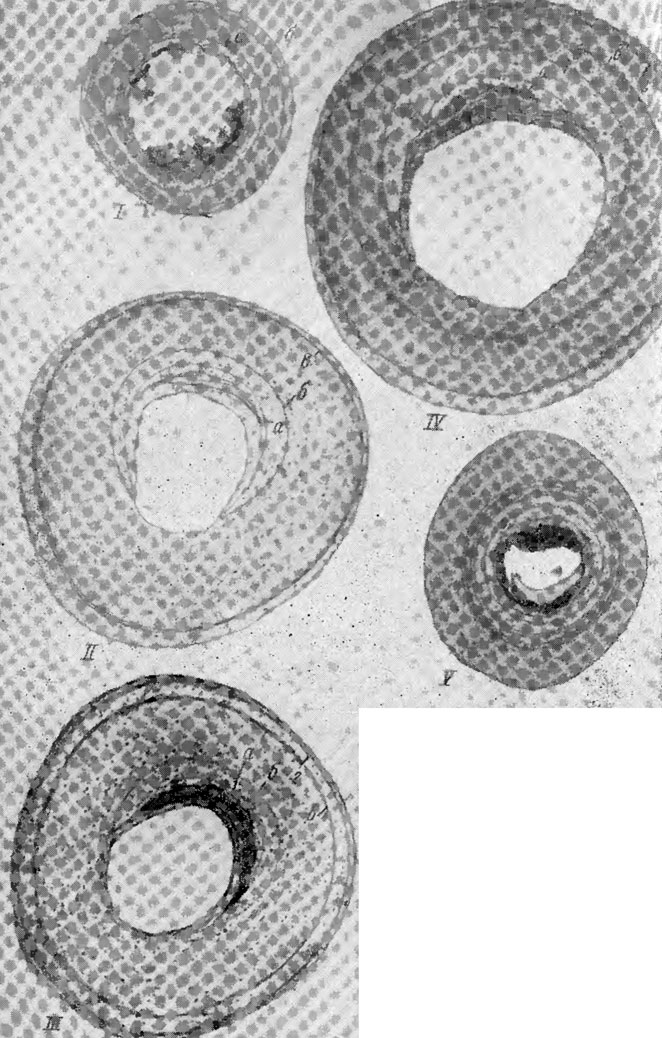
Рис. 78. Поперечные срезы плечевой и бедренной костей прыткой ящерицы I - первая зимовка; II - вторая зимовка; III - третья зимовка; IV - четвертая зимовка; V - случай образования пяти линий склеивания в течение одного года жизни;, линия склеивания: а - у сеголетков; б - первая зимовка (IV, а); в - вторая зимовка (IV, б); г - третья зимовка (IV, в); д - четвертая зимовка (IV, г) (поперечный срез, декальцинированный препарат, окраска гематоксилином, об. 9хок. 15 (препараты и фото Э. М. Смнрнной)
В исследованной выборке из 28 двухгодовиков у 20 видны линии склеивания от двух зимовок, а у 8 экз. линия склеивания второй зимовки не видна. На срезах костей этих экземпляров видны только обе линии склеивания первого года жизни и затем большой прирост костной ткани. Трудно сказать, является ли это особенностью исследованного среза или это нарушение ритма роста и его записи в костях. Однако наша уверенность в том, что эти ящерицы зимовали два раза, а не один, основана на том, что прирост кости от последней линии склеивания до наружного края кости слишком широк, чтобы считать его сформированным со времени пробуждения из последней спячки до середины мая. Возможно, что эти ящерицы еще не начали активно расти и последняя линия склеивания еще не отделилась от наружного края кости. Об этом свидетельствует тот факт, что у некоторых двухгодовиков с линиями склеивания от обеих зимовок, след от второй зимовки проходит около самого наружного края кости, едва отделяясь от него. Длина тела этих ящериц (65 - 76 мм) также дает основание считать их двухгодовиками (см. рис. 77). Вероятно, несоответствие числа слоев возрасту в данном случае можно считать следствием индивидуальных особенностей ритма роста и его записи в костях.
Далее можно выделить группу ящериц, которых нужно считать зимовавшими три раза. От линии склеивания, которая видна у сеголетков, у них обычно остается лишь небольшой участок, и видны еще три линии (одинарные или расслоенные), которые можно считать соответствующими первой, второй и третьей зимовкам (см. рис. 78, III).
И, наконец, встречались ящерицы, которых можно считать зимовавшими 4 раза. У них линия склеивания, которая видна у сеголетков, резорбируется и лишь иногда может оставаться совсем маленький ее участок. Линия склеивания, соответствующая первой зимовке, у них также частично резорбирована, а далее следуют еще три линии (см. рис. 78, IV).
Так как в нашем распоряжении не было ящериц точно известного возраста, для того, чтобы выяснить, сколько линий склеивания успевает резорбироваться со стороны эндоста при активном росте молодого животного, сделали следующие промеры на поперечном срезе кости: у годовиков измеряли диаметр круга, ограниченного первой линией склеивания (которая видна у сеголетков), и диаметр круга, ограниченного второй линией склеивания (которая появляется после первой зимовки). У двухгодовиков и старших особей измеряли диаметр костномозговой полости. Промеры брали у 10 экз. каждой возрастной группы (по три среза от каждой особи). Результаты приведены в табл. 41.
Таблица 41
| Годовики | Двухгодовики | Трехгодовики и старше | |
| Диаметр* кости до первой линии склеивания | Диаметр кости до второй, линии склеивания | Диаметр костномозговой полости | |
| 33,4 x 31,2 | 41,2 x 39,4 | 36,6 x 34,5 | 42,6 x 38,5 |
*(Измерялся наибольший и наименьший диаметры, поскольку срез представляет собой скорее эллипс, а не круг.)
Исходя из этих промеров, можно было бы ожидать, что у двухгодовиков полностью резорбируется первая линия склеивания, а вторая, соответствующая первой зимовке, остается. На самом деле, и с первой этого не происходит, так как резорбция идет не равномерно по кругу, а сильнее захватывает одну сторону кости: поэтому и от первой линии у двухгодовиков виден остающийся участок, который может сохраняться даже у особей, зимовавших три раза. Остаток линии склеивания, соответствующий первой зимовке, сохраняется, очевидно, и у четырехлетних, а, возможно, и более старших особей. Иногда эта линия сохраняется даже целиком.
Таким образом, в выборке из Умани из 93 экз., по нашим предварительным данным, оказалось 49 экз., зимовавших один раз, 28 экз. зимовавших два раза, 10 экз. - три раза и 4 - четыре раза (число зимовок 2 экз. трудно определить).
Следует отметить, что на поперечных срезах костей самых старых из пойманных ящериц ширина каждого из слоев, отложенных после первой, второй, третьей и т. д. спячек, уменьшается незначительно, тогда как у большинства более молодых особей слой, отложенный после первой спячки, намного шире, чем слой, отложенный после второй и последующих спячек. Это может быть следствием одной из двух причин: либо линия склеивания, соответствующая первой зимовке, резорбировалась целиком, и первая линия, которую мы видим, соответствует второй зимовке, либо темп роста этих ящериц был медленным и более равномерным в течение всей жизни и вследствие этого ширина слоя, отложенного после первой зимовки уже, чем обычно. Первое вряд ли вероятно, как видно из приведенных выше промеров костномозговой полости. Второе объяснение кажется более вероятным, и, если это так, то относительное количество экземпляров с равномерно расположенными слоями должно возрастать в каждой последующей возрастной группе. Эта тенденция прослеживается даже на нашем небольшом материале. У всех четырех ящериц, которых мы считаем четырехлетними, ширина слоев на срезах в кости различалась незначительно. Из десяти трехлетних у 6 экз. между линиями склеивания, соответствующими первой и второй зимовкам, был большой разрыв, а у трех линий склеивания располагались почти на одинаковом расстоянии друг от друга (один экземпляр оказался сомнительным). Из 14 экз. двухлетних ящериц лишь у двух линии склеивания разделены небольшими промежутками.
В ихтиологической литературе имеются данные о том, что более быстрорастущие рыбы гибнут раньше, чем медленнорастущие. Поэтому у рыб самая старшая возрастная группа иногда представлена только самыми медленнорастущими особями (Richer, 1969). Может быть, что и у прыткой ящерицы до самых старших возрастов доживают лишь особи с более медленным и равномерным темпом роста. Это положение подтверждается и тем, что размеры тела самых старых из определенных нами: ящериц были не самые крупные (см. рис. 76, 77).
Кроме линий склеивания, разграничивающих годовые слои, на срезах часто видны дополнительные линии, которые, как правило, окрашиваются менее ярко, чем основные линии склеивания, соответствующие зимовкам, но иногда малоотличимы от последних и затрудняют определение возраста. Причиной образования таких линий могут быть, очевидно, влияния любых экзогенных или эндогенных факторов: сильное похолодание летом, засуха, болезнь и т. д., т. е. любой фактор, вызывающий остановку роста. У ящериц на 1 - 2-м годах жизни в костях может образовываться до 4 - 5 близко расположенных полос (см. рис.. 78, V). Это можно объяснить тем, что зимой период спячки у молодых ящериц иногда прерывается короткими периодами активности, вследствие чего у них, хотя и незначительно, могут идти процессы роста (Сергеев, 1937; Davis, 1967), а в летний период и остановки роста (например, в течение летней спячки).
Полученный в данном исследовании материал позволяет сделать вывод о том, что у прытких ящериц ежегодно в период активного весенне-летнего роста в трубчатых костях образуется широкий слой костной ткани, отграниченный от последующего слоя одиночной или расслоенной линией склеивания, образующейся в зимний период. Кроме "зимних" линий склеивания, отграничивающих ежегодные приросты кости, имеется еще одна самая первая линия склеивания, образующаяся у сеголеток вскоре после вылупления. Процессы резорбции костной ткани со стороны эндоста идут у прыткой ящерицы медленно. Первая линия склеивания, образующаяся у сеголетков, целиком резорбируется лишь к 3 - 4 годам. Линия, соответствующая первой зимовке, даже у четырехлетних ящериц целиком может не резорбироваться и, возможно, сохраняется всю жизнь.
Исходя из изложенных выше данных, определение возраста, прытких ящериц по слоям в кости можно считать вполне возможным.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'