Основные черты физиологии прыткой ящерицы и других рептилий
Имеющийся материал позволяет с той или иной степенью подробности рассмотреть следующие жизненные процессы и функции прыткой ящерицы и других рептилий: дыхание и газообмен, терморегуляцию, кровообращение, пищеварение, выделение и некоторые особенности обмена веществ.
Дыхание и газообмен
Прыткой ящерице присуще как кожное, так и легочное дыхание. Точных данных по кожному дыханию у этого вида нет, но по аналогии с другими видами ящериц (Fitzgerald, 1957) можно предположить, что значительная часть углекислоты может выделяться через кожу (до 85% "выделения ее через легкие).
Основное значение при вдохе - выдохе имеет работа мышц боковых стенок грудной полости и шейного отдела. У спокойно сидящей ящерицы дно ротовой полости опускается и поднимается с частотой 2 - 4 раза в 1 сек. Эти движения имеют определенное значение для вентиляции передних частей легких, равно как и для постоянного восприятия запахов при помощи обонятельного мешка, располагающегося по ходу проводящих воздух путей. Значительно более редки глубокие дыхательные движения, совершающиеся за счет сокращения мышц стенок грудной полости. В зависимости от активности животного эти движения совершаются с частотой 1 - 2 в 1 сек., но временами между серией глубоких вздохов может наступать пауза в несколько секунд. Число дыханий резко уменьшается в периоды пониженной активности (ночью, в течение летней и зимней спячки). Вероятно, в такие периоды увеличивается относительное значение кожного дыхания.
Как показывают наши эксперименты, у прыткой ящерицы увеличиваются частота и амплитуда дыхания с повышением концентрации CO2 во вдыхаемом воздухе (подобное явление отмечается и у других видов пресмыкающихся). Таким образом, дыхательный центр ящериц можно рассматривать как прототип дыхательного центра высших позвоночных (Smith, 1939).
Проведенные М. Ф. Тертышниковым (1973) эксперименты* показывают, что с повышением температуры тела потребление кислорода увеличивается, а с понижением уменьшается (табл. 16, рис. 54), что характерно для всех пойкилотермных животных, к которым относятся и прыткие ящерицы. Аналогичные данные известны и для других рептилий (Dawson, Bartholomeu, 1958; Dawson, Templeton, 1963; Калашников, 1964; Benedict, 1932, 1938 и др.).
Таблица 16
| Температура, °С | n | M ± m | n | M ± m |
| Прыткая ящерица | Разноцветная ящурка | |||
| 0 | 8 | 3,0 ± 1,0 | 5 | 14,4 ± 5,8 |
| 5 | 9 | 47,8 ± 4,6 | 23 | 72,2 ± 12,0 |
| 10 | 15 | 66,0 ± 28,94 | 14 | 98,5 ± 11,3 |
| 15 | 20 | 84,2 ± 10,43 | 14 | 124,7 ± 11,3 |
| 20 | 37 | 128,5 ± 17,0 | 18 | 195,0 ± 14,1 |
| 25 | 18 | 242,9 ± 11,2 | 14 | 347,0 ± 23,4 |
| 30 | 16 | 532,0 ± 91,97 | 11 | 1102,0 ± 132,7 |
| 31 | 4 | 169,0 ± 27,75 | 5 | 264,0 ± 73,63 |
| 32 | 8 | 260,0 ± 27,0 | 4 | 256,0 ± 98,65 |
| 33 | 8 | 236,0 ± 20,10 | 3 | 340,0 ± 140,6 |
| 34 | 7 | 113,0 ± 40,0 | 4 | 243,0 ± 61,0 |
| 35 | 13 | 127,0 ± 46,0 | 26 | 273,0 ± 13,6 |
| 40 | 3 | 24,3 ± 2,00 | 3 | 138,0 ± 15,18 |
| 47 | - | 0,00 | 4 | 0,00 |
Наиболее низкое потребление кислорода прыткими ящерицами было отмечено нами при 0°. При такой температуре энергетический обмен очень низок, что, несомненно, биологически оправдано, так как удлиняет продолжительность жизни этих животных в отсутствие корма. Повышение количества потребляемого кислорода происходит при подъеме температуры тела до 30°. При ее увеличении с 30 до 35° количество потребляемого кислорода резко падает и находится на более или менее постоянном уровне. Дальнейшее повышение температуры тела приводит к тому, что газообмен прыткой ящерицы прекращается при 41,6 - 42°, а у разноцветной ящурки - при 47 - 47,2° (при этих температурах ящерицы погибают).
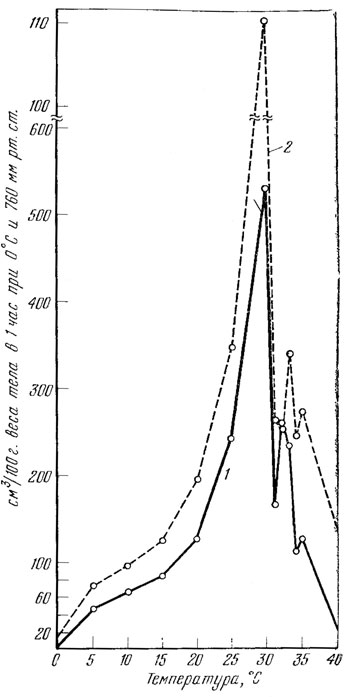
Рис. 54. Характер потребления кислорода прыткими ящерицами (1) и разноцветными ящурками (2) в зависимости от температуры тела (по М. Ф. Тертышникову)
Удержание уровня потребления кислорода в узкой зоне температур от 30 до 35° на более или менее постоянном уровне свидетельствует о наличии у прыткой ящерицы химической терморегуляции. Однако интенсивность этих процессов невысока, что подтверждается их неспособностью удерживать температуру тела на оптимальном уровне (около 30°) при ее снижении или повышении.
Изменение температуры среды на протяжении суток и сезонов сказывается на уровне газообмена. Весной, летом и осенью прослеживается установленная зависимость между количеством потребляемого ящерицами кислорода и температурой. Кроме того, следует отметить, что у прыткой ящерицы уровень газообмена осенью выше, чем летом, а летом выше, чем весной.
Проведенные эксперименты не обнаружили разницы между количеством потребляемого кислорода днем и ночью (при условии, что температурные условия среды не изменялись на протяжении суток).
Между ящерицами разного пола и возраста существуют довольно заметные отличия в уровне газообмена (табл. 17). Возрастные отличия в количестве потребляемого кислорода отмечены и для других групп рептилий (Калашников, 1964). Причину данного явления следует искать, видимо, во взаимосвязи между уровнем окислительных процессов в организме и размерами тела. Более интенсивное поглощение кислорода молодыми животными объясняется относительно большей поверхностью их тела (по отношению к весу), чем у взрослых животных (Rubner, 1883).
Таблица 17
| Группа особей по 5 экз. | Количество кислорода, см3 | Температура тела, °С | Вес животного, г |
| Взрослые самцы | 1410 | 11,5 - 25 | 13,0 - 25,0 |
| Взрослые самки | 1063 | 14,0 - 19,5 | 13,0 - 20,0 |
| Полувзрослые самцы и самки | 3330 | 17,0 - 24,0 | 1,4 - 2,8 |
Терморегуляция
Обычно наибольшая активность прытких ящериц наблюдается при размахе предпочитаемых температур тела 18,0 - 36,0° (измерения в клоаке), причем в разных географических районах температура наибольшей активности несколько различна и, видимо, является локальной адаптацией к конкретным условиям существования (Тертышников, 1973а; Боченко, уст. сообщ. и др.). В среднем максимальная активность наблюдается при температуре тела, близкой к 30° (рис. 55). При температуре тела ниже 7,5° прыткая ящерица впадает в оцепенение, при температуре тела ниже -4,9° и выше 42° погибает (Либерман, Покровская, 1943; Стрельников, 1944).
Для прыткой ящерицы характерны значительные суточные и сезонные колебания температуры тела. Данные о топографии температуры тела отсутствуют, однако прямые наблюдения в природе и террариуме показывают, что температура конечностей и хвоста обычно заметно ниже температуры брюха и горла.
Итак, несомненно, что температура тела ящерицы может значительно отличаться от температуры окружающей среды. По наблюдениям М. Ф. Тертышникова, эти различия достигают 18°. А. В. Яблоков и А. В. Валецкий в апреле 1973 г. в районе г. Караганды наблюдали прытких ящериц, активно бегающих по снегу в солнечную ветреную погоду при температуре воздуха до -10°; в этом случае разница между температурой среды и температурой тела должна была быть не менее 25 - 30° (!). Поэтому, строго говоря, отнесение этого вида к пойкилотермным животным, у которых отсутствует терморегуляция, может быть сделано лишь с большими оговорками. Видимо, правильнее считать этот вид ящериц (как видимо и большинство других рептилий) гетеротермным. Пока мало известно о механизмах химической терморегуляции данного вида, хотя сам факт такой терморегуляции несомненно имеет место (М. В. Тертышников). Значительно больше данных имеется по физической терморегуляции, которая у прыткой ящерицы осуществляется всеми тремя известными способами - проведением, излучением и испарением - (Либерман, Покровская, 1943; Стрельников, 1944; Тертышников, 1973 и др.) и тесно связана с поведенческой активностью (см. гл. X).
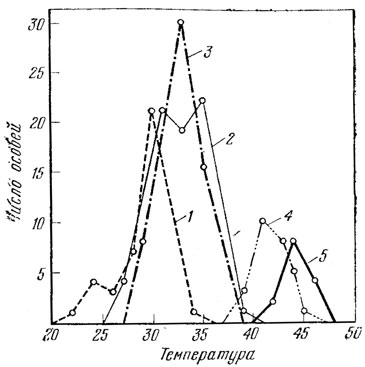
Рис. 55. Основные температурные параметры прыткой ящерицы: 1 - температура активных ящериц в природе; 2 - температура, избираемая ящерицами в эксперименте в аппарате Гертера; 3 - температура тела ящериц при температуре воздуха 32,8° С (оптимальной); 4 - температура теплового окоченения прыткой ящерицы; 5 - летальная температура. Число исследованных особей соответственно: 37, 65, 79, 21 и 14 экз. (по данным С. С. Либермана, Н. В. Покровской, 1943)
Поскольку прыткая ящерица покрыта прочной роговой чешуей, до последнего времени считалось, что животные теряют мало воды путем испарения. В связи с этим предполагалось, что температура тела прыткой ящерицы будет мало зависеть от влажности. В пользу этого суждения свидетельствует тот факт, что у жабровидной ящерицы снижение влажности со 100 до 7% при 20° вызывало понижение температуры тела всего на 1° (Hall, Root, 1930).
Однако в последние десятилетия накопилось немало данных, показывающих, что чешуйчатый покров ящериц не является препятствием для испарения влаги организмом. Прямые эксперименты показали, что через кожу ящерицы могут испарять от 25 до 50% всей теряемой организмом влаги, и кожа является таким образом важным органом активной физической терморегуляции (Claussen, 1967; Kerfoot, 1970 и др.). Об этом же косвенно свидетельствует высокая положительная корреляция числа чешуй у некоторых видов североамериканских ящериц с минимальными зимними температурами, с одной стороны, и величиной осадков в летние месяцы с другой (Soule, Kerfoot, 1972). К этому остается добавить, что существует клинальная изменчивость числа чешуй у некоторых вbдов рептилий, что также свидетельствует о возможной роли чешуйчатого покрова в терморегуляции.
Итак, температура тела ящериц во многом зависит от температуры внешней среды, однако прыткая ящерица имеет некоторые механизмы, регулирующие температуру. Эта регуляция у прыткой ящерицы достигается изменением как поведения (см. гл. X), так и метаболизма.
Характеристика крови
У прыткой ящерицы содержание гемоглобина составляет 7,8 г на 100 мл крови (Тертышников, 1972). О продолжительности жизни эритроцитов крови у прыткой ящерицы сведения отсутствуют, но можно предположить, что она равна 11 месяцам, как у большинства других .пресмыкающихся (Berlin, 1959).
По нашим данным, количество эритроцитов у прыткой ящерицы составляет 11 670 в мм3 крови, РОЭ -4 мм в час, гемоглобин - 11,5%. У близких видов ящериц полосы поглощения различных форм пигмента крови характеризуются такими величинами: HbO2 - 576,2 ммк, HbCO - 571,5, расстояние между α-полосами HbO2 и HbCO - 4,7 ммк (Prosser, Brown, 1962). Нет данных по прыткой ящерице о видах гемоглобина, которые у пресмыкающихся отличаются по молекулярному весу, изоэлектрической точке и времени денатурации (Ramirez, Dessauer, 1957). Константа седиментации гемоглобина у L. vivipara равна 4,6 S. Поскольку эта константа свойственна рыбам, пресмыкающимся, птицам и млекопитающим (Svedberg, Pedersen, 1959), то и прыткая ящерица, видимо, не должна составлять исключения.
Дыхательные свойства крови у настоящих ящериц, к которым относится и L. agilis, характеризуются следующими величинами: кислородная емкость крови составляет 10 - 12,5 об.%, напряжение O2 при 50% насыщения крови (P50) равно 19 - 31 мм рт. ст. (Edwards, Dill, 1935; Dawson, Bartholomew, 1960).
Касаясь взаимосвязи дыхательных пигментов у ящериц с экологическими условиями следует сказать, что, чем доступнее кислород для организма, тем выше давление, при котором происходит насыщение гемоглобина кислородом. У прыткой ящерицы, по нашим данным, не отмечено усиления эритропоэза при низком парциальном давлении кислорода. При этом у прыткой ящерицы имеет место лишь усиление дыхания. О переносе углекислого газа у ящериц известно лишь то, что его содержание в крови колеблется от 38,6 до 44 об. % (Hopping, 1923).
Таблица 18
| Популяция (УССР) | Альбумины | Глобулины | |||
| 1 | 2 | α1 | β1 | ν1 | |
| Равнинная (Подолия) | 36,699 | - | 43,615 | 4,265 | 15,417 |
| Субальпийская (Карпаты) | 39,062 | 33,854 | 12,500 | 3,645 | 10,937 |
У ящериц сывороточные белки крови имеют хорошо выраженные буферные свойства (Redfield, 1933, 1934). У прыткой ящерицы обнаружено несколько фракций крови. Данные о составе белковых фракций сыворотки крови для двух западных популяций прыткой ящерицы приведены в табл. 18, а денсито-метрическая кривая - на рис. 56.
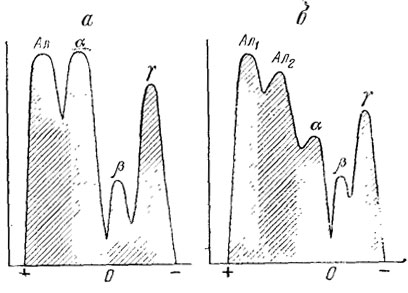
Рис. 56. Денситограмма сыворотки крови прыткой ящерицы: а - равнинная популяция в Подолии; б - горная популяция на альпийском лугу в Карпатах (Страутман и др., 1965)
Из приведенных данных видно, что характер фракций крови у прыткой ящерицы значительно колеблется в разных популяциях (Черкащенко, Кушнирук, 1973).
Относительный объем крови у пресмыкающихся сильно варьирует (Coulson, 1950; Semple, 1960). По нашим наблюдениям, у прыткой ящерицы объем крови составляет 6,3% веса тела.
Кровообращение
Как отмечено в главе III, вес сердца у прыткой ящерицы составляет 3,4 - 3,9% веса тела, что очень близко к показателям у других представителей этого рода (Quiring, 1950). Наши наблюдения показывают, что число сокращений сердца при 20° у взрослых ящериц составляет 52 ± 2 ударов/мин. Другие авторы (Laurens, 1920) приводят несколько меньшие величины, которые, однако, даже еще меньше, чем у других представителей этого рода (Buchanan, 1909). При этом надо отметить, что изменение температуры среды на 1° в диапазоне 18 - 32° вызывает у прыткой ящерицы изменение частоты сокращения сердца в среднем на 1 удар в минуту.
Данных по кровяному давлению в малом и большом кругах кровообращения у прыткой ящерины нет, однако, по всей вероятности, они должны быть близки к этим показателям у других видов ящериц. У серого варана (Varanus griseus), например, по нашим данным, кровяное давление в левом "желудочке" сердца равно 65 - 75/0, а в дуге аорты - 65 - 75/30 мм рт. ст. В малом круге кровообращения уровни кровяного давления характеризуются такими величинами: в правом "желудочке" сердца - 42 ± 1/0, а в легочной артерии - 41 ± 1/22 ± 1 мм рт. ст. Эти показатели близки к таковым у других видов ящериц (Harrison, 1965; White, 1968). Как показывают наши эксперименты, минутный объем сердца на килограмм живого веса у серого варана равен 112 мл, общее сосудистое сопротивление большого круга кровообращения составляет 19 980, а малого круга кровообращения - 8880 дин/сек/см-5.
Известно, что у пресмыкающихся повышение температуры тела ведет к повышению кровяного давления. Таким образом, у ящериц, вероятно, существуют вазомоторные механизмы экзотермической терморегуляции, когда расширение сосудов способствует поглощению тепла извне, а сужение их препятствует этому явлению (Prosser, Brown, 1962).
Несмотря на то, что у ящериц межжелудочковая перегородка неполная (Foxon, 1955; Татаринов, 1960; Жеденов, 1961; Ludicke, 1964; Антипчук, 1966, 1967; Антипчук, Гибрадзе, 1973), существ вует мнение, что потоки крови в трехкамерном сердце ящериц в какой-то мере разделены (Hoffman, 1879 - 1890; Соре, 1900; Goodrich, 1930; Шмальгаузен, 1938; Meinertz, 1952, 1966; Foxon, 1956; Татаринов, 1960; Жеденов, 1961; Ludicke, 1964). С одной стороны, физиологические данные свидетельствуют о том, что у ящериц существует незначительное смешивание крови (Prakash, 1952; Foxon et all., 1953; Foxon, 1955; Khalil, Zaki, 1964; Grasse, Devillers, 1965) в трехкамерном сердце. С другой стороны, в противоположность приведенным данным,, существует мнение, что при оптимальной температуре тела у ящериц имеет место полное разделение артериальной и венозной крови в трехкамерном сердце (White, 1955, 1956, 1959, 1968; Антипчук, 1966, 1967). Объяснение подобного противоречия приводится в книге А. П. Антипчука, Т. А. Гибрадзе (1973). Поскольку у прыткой ящерицы сердце трехкамерное, то и у нее при оптимальных температурах должно иметь место полное разделение потоков артериальной и венозной крови.
Пищеварение
Известно, что способ питания данного организма определяется главным образом средой обитания и характером доступной пищи. Гистохимическое исследование двенадцатиперстной кишки у 33 видов ящериц (в том числе и у прыткой ящерицы) показало, что строгая зависимость между характером питания и гистоструктурой отсутствует (Gabe, Girons, 1972).
Касаясь переваривания углеводов, можно сказать, что у ящериц обнаружена в кишечнике активная мальтоза. Поскольку мальтоза и сахароза, как правило, встречаются вместе и оптимумы их рН близки, можно предположить, что мальтоза и сахароза перевариваются при помощи одного и того же фермента (Yonge, 1928; Manseur-Bek, 1932, 1933, 1948). Кроме того, имеются данные, показывающие, что у прыткой ящерицы лактоза в кишечнике не переваривается. По нашим данным, у взрослых прытких ящериц химус желудка имеет кислое рН, и в нем обнаружен пепсин.
О переваривании жиров у прыткой ящерицы прямых данных нет, однако известно, что у большинства рептилий в кишечнике имеется триоксикопростановая и питохолевая кислоты (Bergmann, Расе, 1943). Эти кислоты также должны быть присущи и прыткой ящерице, у которой, как и у прочих рептилий, активность пищеварительного сока связана с панкреатической липазой. Однако у ящериц не установлено, какой фермент принимает участие в переваривании жиров - липаза или эстераза. Состав потребляемого жира у ящериц до некоторой степени определяет состав депонированного жира, в который входит значительное количество ненасыщенных жиров. Депонированный жир у ящериц содержит много олеиновой кислоты и меньше пальмитиновой и стеариновой кислот (Klenk, 1933, 1935; Крепе, 1964).
Данные по скорости переваривания пищи, количественному и качественному составу пищи в разных частях ареала приведены в главе VI.
Выделение
У ящериц, как и у других пресмыкающихся, конечные продукты обмена веществ удаляются из организма путем диффузии и активных процессов выделения. Аминокислоты подвергаются окислительному или гидролитическому дезаминированию с выделением свободного аммиака. По нашим данным, у прыткой ящерицы концентрация аммиака в крови достигает 0,12 мг/100 мл, т. е. примерно такая же, как и у других видов пресмыкающихся. Ящерицы способны превращать аммиак в мочевую кислоту. Последняя мало токсична и плохо растворима; она может откладываться или выделяться в кристаллической форме. Точка замерзания мочи у прыткой ящерицы, по нашим наблюдениям, 0,2, а крови 0,62, т. е. находится в пределах, свойственных другим пресмыкающимся (Smith, 1932; Bentley, 1959).
Непосредственным источником аммиака у ящериц является глютаминовая кислота, а источником выделяемого азота - продукты обмена нуклеиновых кислот - пурины. Азот пиримидинов выделяется у ящериц в виде мочевины и аммиака, причем мочевая кислота образуется как конечный продукт окислительного дезаминирования пуринов. Птерины насекомых, которыми питаются ящерицы, входят в состав фолиевой кислоты - птероил-L-глютаминовой кислоты. Биосинтез птеринов во многих чертах напоминает биосинтез мочевины. Кроме того, существует связь между биосинтезом птеринов и биосинтезом рибофлавина. Эти вещества встречаются в хроматофорах пресмыкающихся (Forrest, et al., 1962).
У большинства рептилий процентное соотношение конечных продуктов азотистого обмена (по азоту) составляет: аммиака - 8,7, аминного азота - 2,3 и мочевой кислоты - 89% (Bacon, 1909). Следовательно, у ящериц большая часть азота выделяется в виде мочевой кислоты (Khalil, Haggag, 1955) и, вероятно, прыткая ящерица не составляет исключения; поэтому ее можно отнести к уриколитическим животным. Различие между уриколитическими и уролитическими организмами является чисто количественным, и наличие мочевой кислоты в крови еще совсем не означает, что мочевая кислота является основным продуктом азотистого обмена.
Известно, что у чешуйчатых рептилий катаболизм белков изменяется на различных стадиях онтогенеза. На ранних стадиях развития конечные продукты белкового обмена представлены аммиаком: далее возрастает удельный вес мочевины и затем большая часть азота выделяется в виде мочевой кислоты, скапливающейся в аллантоисе. Кроме того, у чешуйчатых в период уменьшения выделения мочевины в аллантоисе во время развития зародыша обнаруживалась уреаза в печени и почках (Clark, 1953). Подобное изменение в составе конечных продуктов азотистого обмена в раннем онтогенезе у ящериц биологически оправдано, так как токсичность азотистых продуктов обмена снижается в направлении от аммиака к мочевой кислоте.
Для развития яиц ящерицы большое значение имеет процесс задержки расходования воды. В связи с этим в развивающихся яйцах ящериц продукты азотистого обмена сохраняются до вылупления и, как показывают наши данные, эти продукты у прыткой ящерицы представлены в основном мочевой кислотой.
Известно, что у пресмыкающихся отщепление аммиака от аминокислот катализируется дезаминазами. Мочевина образуется в орнитиновом цикле при действии аргиназы на аргинин и аллантоиказы на аллантоиновую кислоту. В пользу этого суждения свидетельствует тот факт, что у чешуйчатых в почках обнаружена аргиназа, расщепляющая экзогенный аргинин (Clark, 1953; Clark, Sisken, 1956; Goldie, 1959). Возможно, что у чешуйчатых мочевая кислота синтезируется при участии карбамилфосфата, а источниками аммиака являются глютаминовая и аспаргиновая кислоты. При этом требуется участие кислот трикарбонового цикла и энергия АТФ. Кроме того, синтез мочевины возможен путем расщепления пуринов ксантинокиназой (Prosser, Brown, 1962).
Поскольку у ящериц из ферментов, расщепляющих мочевую кислоту, обнаружена лишь ксантиноксидаза, у них пурины могут дезаминироваться до ксантина или гипоксантина, которые затем превращаются в мочевую кислоту (Florkin, Duchateau, 1943; Laskowski, 1951; Albritton, 1954). По всей вероятности, прыткая ящерица в этом отношении не является исключением.
Данные о функции почек у ящериц не многочисленны. Можно отметить в этой связи, что почки бедны кровеносными сосудами. Нет сведений о резорбции воды в мочевом пузыре. Считается, что почки лишь частично регулируют экскрецию воды и электролитов. По современным представлениям, у ящериц резорбция большей части воды, имеющейся в моче, происходит в клоаке (Green, 1972). Наши наблюдения показывают, что клоакальная моча прыткой ящерицы может быть вязкой вследствие наличия кристаллов мочевой кислоты. Количество мочи, выделяемой прыткой ящерицей, составляет 5,1 мл/час на кг веса тела, т. е. примерно так же, как и у прочих видов ящериц (Bentley, 1959).
Органы чувств
В последнее время появились специальные исследования органов чувств у прыткой ящерицы (Swiezawska, 1949; Dudziak, 1955; Viand, 1960; Васильев, 1973). Однако работа органов чувств данного вида все же остается далеко не полно изученной.
Слух. Имеются данные (без указания вида), что ящерицы способны воспринимать звуки в диапазоне всего лишь 7,4 - 8,2 кгц (Buddenbrock, 1952; по Никитенко, 1969); эти данные нуждаются в проверке на более совершенном современном оборудовании. Известно, что прыткая ящерица иногда шипит, с силой выпуская струю воздуха сквозь суженный просвет глотки. Этот звук, несомненно, воспринимается другими особями как сигнал угрозы. Недавно описаны (Баранов; и др., 1972) звуки прытких ящериц, напоминающие пение, однако биологическое значение их остается неясным (звуковая активность в условиях этих наблюдений была, по всей видимости, связана с половой активностью). Не исключена возможность того, что при тщательном специальном исследовании окажется, что ящерицы способны к восприятию ультразвуковой и инфразвуковой сигнализаций. Инфразвуки, хорошо передающиеся через твердый грунт на значительное расстояние, несомненно должны улавливаться через участки тела, прилегающие к грунту.
Известно, что многие виды ящериц реагируют на звуки в диапазоне частот от 8000 до 10 000 гц, однако нет никаких данных о том, что они способны различать звуки различной частоты (Adrian, 1938, 1943; Wever, Vernon, 1956).
Исследование чувствительности слухового анализатора у прыткой ящерицы путем отведения биопотенциалов от слуховых ядер продолговатого мозга показало, что они обладают хорошей чувствительностью к высоким звукам. Низкие звуки воспринимаются прыткими ящерицами в диапазоне частот 200 - 6000 гц. При этом ящерицы воспринимают разнокачественные сигналы с четкой дифференцировкой и изменение амплитуды ответной реакции на щелчок происходит скачкообразно (с быстрым переходом к насыщению), а изменение амплитуды ответов на тоны меняющейся интенсивности носит почти линейный характер (Васильев, 1973). Способность медуллярных ядер продолговатого мозга воспринимать повторные раздражения на 99% восстанавливается у прыткой ящерицы уже через 60 мсек. Увеличение фронта нарастания амплитуды стимулирующего сигнала у прыткой ящерицы приводит к падению амплитуды ответа, хотя и при различных значениях экспоненты (Васильев, 1973).
Зрение. Строение зрительного анализатора (см. главу III) свидетельствует о том, что для прыткой ящерицы должна быть характерна чувствительность к красному, оранжевому и желто-зеленому цветам (Forbes a. oth., 1959). Экспериментально показано, что прыткая ящерица может различать от 5 до 8 цветов (Viand, 1960). Вывод о развитии цветового восприятия у прыткой ящерицы подтверждается косвенно и яркой окраской наружных покровов. Язык зрительных сигналов у многих пресмыкающихся основан на смене и показе в определенной последовательности специфически окрашенных участков тела, поз и движений. Если основываться на наблюдениях Г. Китцлера (Kitzler, 1941), который показал, что закрашивание в синий цвет горла у самки зеленой ящерицы (Lacerta viridis) приводит к тому, что на нее нападают самцы (сине-голубое горло является вторичным половым признаком самца), то вполне понятным становится такое же поведение самцов прыткой ящерицы, нападающих на самок, тело или горло которых окрашены в зеленый цвет. Острота зрения прыткой ящерицы была определена в эксперименте, "проведенном Я. Дудчеком (Dudziak, 1955). Ящерицы различали расстояние между предъявляемыми им полосами в 1,8 мм с дистанции 15 см.
Другие органы чувств. Данных по функционированию других органов чувств у прыткой ящерицы практически нет. Некоторые заключения, основанные на наблюдениях в природе и анализе строения специальных структур, касающиеся работы органа осязания, обоняния и теменного глаза, приведены выше (см. главу III).
О физиологии нервно-мышечного аппарата
Прямых данных о физиологии нервно-мышечного аппарата у прыткой ящерицы не имеется, однако процесс нервно-мышечного проведения изучен на ящерицах других видов (Eyzdguirre, 1957). У некоторых пресмыкающихся установлена зависимость между скоростью проведения нервных импульсов и диаметром нервного волокна (Tasaki, 1959). Как показывают наши наблюдения, мышцы легких прыткой ящерицы способны к длительному спонтанному сокращению, что свойственно также ящерицам других видов (Serfaty, Peyrand, 1960). У некоторых пресмыкающихся показано существование центров вдоха и выдоха, но не установлено наличия пневмотоксического (тонического) центра. У ящериц с удалением мозжечка уменьшается спонтанность движения и изменяется походка, что свидетельствует об участии мозжечка в процессе координации движений (Prosser, Brown, 1962).
У ящериц локальное раздражение переднего мозга может вызывать моторные реакции (Prechtl, 1949), а раздражение среднего мозга усиливает электрическую активность переднего мозга и вызывает судорожные реакции (Winkel, Caspers, 1953).
Физиология желез внутренней секреции
Наши знания об эндокринных регуляциях пресмыкающихся пока еще скудны. Ниже приводятся отрывочные данные по этому разделу физиологии рептилий, которые позволяют в общей форме представить основные черты этих процессов и у прыткой ящерицы.
Известно, что у чешуйчатых рептилий длительное кормление щитовидной железой ведет к повышению интенсивности обмена веществ, повышает возбудимость и ведет к значительной потере веса ( Krockert, 1941).
Чувствительность тканей пресмыкающихся к инсулину очень низкая (Sturkie, 1954; Miller, Wurster, 1956), а удаление поджелудочной железы ведет у них к развитию гипергликемии (Foglia, et all., 1955). Вместе с тем пресмыкающиеся весьма чувствительны к глюкагону, их поджелудочная железа особенно богата а-клетками, которые и продуцируют глюкагон (Houssay, 1959). У прыткой ящерицы 'кормление в эксперименте вызывало гликемию и увеличение содержания глюкогена в печени (Vladescu, et all., 1971). Введение инсулина при этом приводило к развитию гипогликемии.
У пресмыкающихся, как у прочих пойкилотермных животных, нейрогипофизарные гормоны характеризуются тем, что их действующим началом является вещество, близко стоящее к вазопрессину, а именно - аргинин-вазопрессин (Heller, 1942; Ewer, 1952; Sawyer, 1955, 1960; Sawyer, Schisgall, 1956; Sawyer, et all., 1959, 1960).
Как и у прочих пойкилотермных животных, очевидно, у них имеется меланоцитостимулирующий гормон, который продуцируется аденогипофизом. Этот гормон вызывает рассеивание пигмента в меланофорах животных, что ведет к сезонному изменению тональности их окраски (Leloup, Berg 1964; Geschwind et all., 1956, 1957; Lee, Lerner, 1957; Harris, Roos, 1956; Harris, Lerner, 1957; Gilbert, Schneiderman, 1958). При этом на изменение окраски оказывает влияние кроме гормональной также и нервная системе (Redfield, 1918; Slome, Hogben, 1928; Parker, 1940; Matsumoto, 1954). Удаление гипофиза у ящериц приводит к посветлению кожных покровов. Хроматофоры ящериц могут принимать участие в терморегуляции. Кроме того, существуют данные о регуляции изменения формы меланофоров при участии вегетативной нервной системы (Parker, 1906; Bauer, 1914; Kruger, Kern, 1924). У пресмыкающихся при повышении температуры среды происходит концентрация меланина в меланоцитах, а при понижении - его дисперсия. По всей вероятности, это служит регулятором света, поглощаемого меланином (Parker, 1940). Все изложенное должно быть справедливо и для прыткой ящерицы.
Существует мнение, что пресмыкающимся свойственны гона- дотропные, фолликулостимулирующие, лютеинизирующие и лю- теотропные гормоны (Hisaw, 1959). Доказано, что функция половых желез и развитие вторичных половых признаков у пресмыкающихся регулируется гормонами гипофиза (Combescot, 1955). У ящериц при введении фолликулостимулирующего и лютеотропного гормонов наблюдается увеличение яичников (Mellish, Meyer, 1937). Андрогены и эстрогены обнаружены в крови ящериц (Bragdon et all., 1954; Amoroso, 1955; Matthews, 1955; Panidel, 1956). Все сказанное присуще, очевидно, и прыткой ящерице.
В регуляции основных физиологических процессов в организме ящериц важную роль играют жировые тела (Hann, Tinkle, 1965). Во время спячки и размножения биохимический состав последних заметно изменяется и, в частности, резко колеблется содержание жирных кислот, холестерола, фосфолипидов, гликогедена и каротиноидов (Hadley, Christie, 1974). Динамика содержания липидов в развивающихся яйцах прыткой ящерицы описана в работе 3. Гродзинского (Grodzinski, 1949).
Из всех известных внешних раздражителей, влияющих на репродукцию, наиболее действенным, очевидно, является свет (Rowan, 1938; Bartholomew, 1949, 1956; Jenner, Engels, 1959; Farner, 1955, 1959; Wolfson, 1959). Прыткой ящерице, как и всем остальным видам ящериц, присущи резкие фотопериодические изменения полового цикла. Об этом свидетельствуют как наблюдения в природе, так и экспериментальные данные. По наблюдениям А. Б. Стрельцова, отловленные в июне - августе и содержащиеся в террариуме ящерицы к ноябрю стали вялыми, перестали питаться, по бокам появились складки кожи, свидетельствующие об истощении. Ящерицы начали прятаться под камни, закапываться в землю, хотя температура под электролампой в террариуме достигала 60°.
После установления ультрафиолетовой лампы (ПРК-8 на тлеющем разряде ежедневно 5 час. облучения) через две недели активность ящериц резко возросла. Они интенсивно передвигались по террариуму, нормально питались, началась массовая линька. Самцы, окраска которых после отлова постепенно бледнела, становясь все более желто-коричневой, после облучения стали зеленеть и приобрели ярко-зеленую окраску, типичную для многих самцов в разгар половой активности. Наконец, 27 декабря наблюдалось первое спаривание.
У некоторых ящериц гормон щитовидной железы стимулировал в экспериментах линьку (Maderson and oth., 1970). Интересно, что у змей тиреоидные гормоны не оказывали такого действия.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'