Группа rudis
Lacerta rudis rudis Bedriaga, 1886
(Рис. 41; фот. 14)
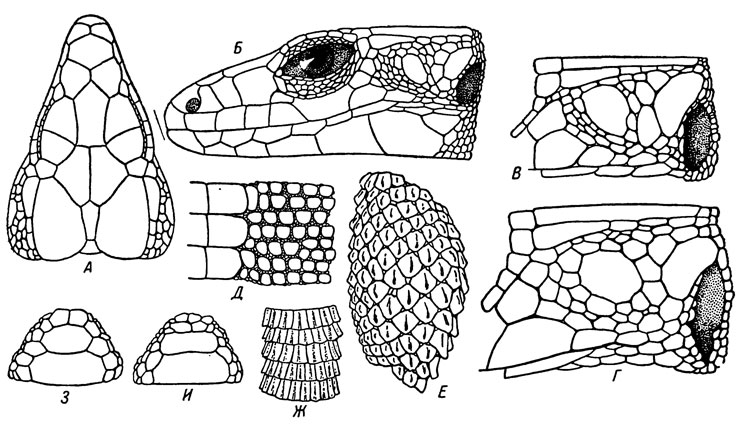
Рис. 41. Основные признаки фолидоза L. r. rudis: А - голова сверху; Б - голова сбоку; В, Г - височная область; Д - пограничная область между туловищной чешуей и щитками брюха самки; Е - верхняя сторона голени; Ж - верхняя сторона передней трети хвоста; З, И - анальная область. Г-Ж - Местия, остальпые - Аджария)
Podarcis depressa Camerano (part.), 1878 : 539. - Lacerta depressa var. rudis Bedriaga, 1886:275 (259). - muralis var. depressa f. modesta, Boettger (part.), 1892:140. - muralis var. depressa, Boulenger, 1904:337, tab. XXII, fig. c; 1913:196, tab. XXII, fig. 7, 7a, 8. - muralis var. rudis Boulenger, 1920 : 290. - saxicola rudis, Mehely, 1909:529, tab. 20, fig. 7; Никольский, 1915:373; Lantz et Cyren, 1936:165; Терентьев и Чернов, 1949:187.

Фот. 14. L. r. rudis (А-В - Верхняя Сванетия, Г-Д - Аджария)
Голотип. Неизвестен. Описана Бедрягой (Bedriaga, 1886) по экземплярам из Батуми.
Описание. Ширина лобоносового превышает, реже равна его длине. Межчелюстной отделен от лобоносового или изредка соприкаса ется с ним в одной точке. Шов между лобоносовым и задненосовым не короче шва между передним и задним носовыми. Швы между предлобными и лобным прямые или слабо вогнуты внутрь лобного. Верхнерес ничные отделены от надглазничных полным или в той или иной мере прерванным рядом из 13-25 зернышек. Верхний заглазничный обычно более или менее широким швом касается теменного. Первый верхневи сочный длинный или умеренно длинный, заметно суженный сзади. Позади верхневисочного на границе с теменным расположено 2-6 мелких, слабо выраженных задневисочных. Большой или средней величины централь новисочный отделен от верхневисочного 2-4, а от барабанного умеренной величины 1-5 продольными рядами мелких щитков. У взрослых особей, особенно у самцов, мелкие щитки височной области нередко несут на себе короткие притупленные шипики или тупые продольные ребрышки.
Воротник не зазубрен. По средней линии горла до воротника 20-32 чешуйки. Чешуя туловища выпуклая, заметно увеличенная на боках, как правило, с выраженными в той или иной мере продольными ребрышками или коническими бугорками, всегда более четкими в задней трети спины и на границе с брюхом. Вокруг середины тела в одном ряду 38-53 чешуйки. Брюшные щитки соприкасаются по бокам тела с 3, реже с 2 туловищными чешуйками у самцов и 2, реже 3 у самок; соответственно брюшные и грудные щитки располагаются в 21-25 поперечных рядов у первых и 24-28 у вторых. Анальный большой, впереди него симметрично расположены один крупный или, реже, два или три более мелких преанальных. Бедренных пор 13-25. На нижней стороне бедер между бедренными порами и наружным краем увеличенных чешуй расположено 4-6 поперечных рядов мелких щитков. Чешуя, покрывающая голень сверху, значительно крупнее спинной, с хорошо развитыми продольными ребрышками и обычно мелкими зернышками по углам; средние чешуйки, как правило, заметно крупнее краевых. Вокруг середины голени в одном ряду 10-15 чешуй. Верхние и боковые чешуи хвоста с сильно развитыми продольными ребрышками, тупо обрезанные сзади или вытянутые более или менее выраженным углом назад; в верхней трети хвоста концы ребрышек заметно вздернуты и образуют острые шипики, вследствие чего хвост взрослых особей выглядит шиповатым. На нижней стороне хвоста чешуйки со сглаженными, но вполне заметными продольными ребрышками. Длина туловища с головой 63-85 мм у самцов и 54-83 мм у самок; отношение ее к длине невосстановленного хвоста соответственно равно 0.48-0.60 у первых и 0.47-0.63 у вторых.
Окраска верхней стороны тела самцов зеленая, светло-зеленая, зеленая как плющ, оливково-зеленая, буровато-зеленая, темно-песочная, бледнобурая или бурая; самок - темно-песочная, оливково-серая, темно-мышино-серая или ореховая. Центральная затылочная полоса образована сосредоточенными посредине спины крупными вытянутыми поперек черными или темно-бурыми пятнами и ограничена по сторонам более или менее широкими, лишенными рисунка полосками цвета основного фона, иногда с четкими светлыми рябинами. Височные полосы образованы обычно тремя рядами более или менее четко выраженных, соприкасающихся друг с другом черных или бурых кружков с хорошо заметными беловатыми (голубыми на уровне передних ног) центрами. Верхние края этих полос ограничены обычно четкими беловатыми глазками, слагающими пунктирные височные линии. Брюхо и нижняя сторона головы и горло самцов зеленовато-желтое, желтоватое, сизо-зеленоватое, голубоватое или голубое. Самки снизу желтоватые или сизо-зеленоватые. В период размножения крайние брюшные щитки самцов и пограничные с ними участки тела ярко-синие или фиолетовые.
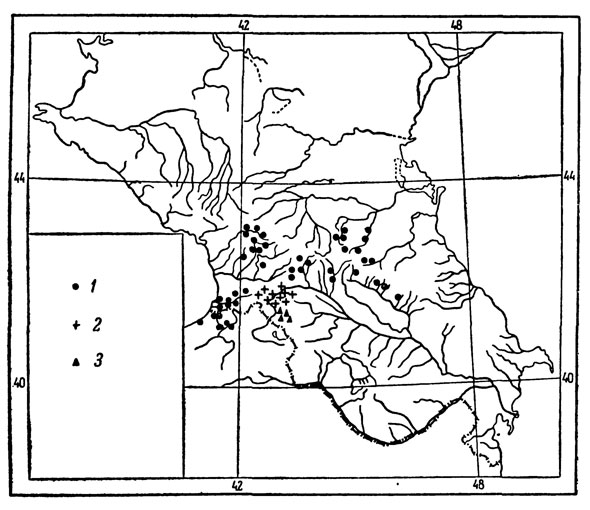
Рис. 42. Основные местонахождения на Кавказе и в северо- восточной Турции: 1 - L. r. rudis; 2 - L. r. obscura, 3 - L. r. macromaculata
Географическое распространение. Обладает широким ареалом, встречаясь в северо-восточной Турции и Западном Закавказье, с одной стороны, и на Большом Кавказе в пределах северной Грузии, северо-западного Азербайджана и горной Чечено-Ингушетии - с другой. Юго-западная граница ареала проходит в Малой Азии по северным склонам Понтийского хребта где на севере распространение подвида ограничено береговой линией, а на востоке захватывает долину нижнего течения р. Чороха и его притоков. В Аджарии обычна в долине р. Аджарис-Цкали и ее притоков, берущих начало на склонах Месхетского, Шавшетского и Арсиянского хребтов. По северным предгорьям Месхетского и далее по Сурамскому хребту достигает южных предгорий Большого Кавказа, где широко распространена в пределах северной Грузии, южной Осетии и северо-западного Азербайджана - от долины среднего и верхнего течения р. Ингури в Мингрелии и Сванетии на западе до окр. Закатал на востоке. Южная граница ареала на Главном хребте проходит по предгорьям Эгрисского, Лечхумского и Рачинского хребтов до Накалакеви и Кутаиси на юге, затем по предгорьям хребтов, ограничивающих с севера Внутреннекартлинскую равнину, и, пересекая Картлинский и Кахетинский хребты, продолжается далее по южным предгорьям Большого хребта в долине Алазани до Закатальского заповедника в пределах Азербайджана. Обширная изолированная популяция существует на северных склонах Большого хребта в Чечено-Ингушетии, откуда эта ящерица впервые была указана Черновым (1929) из ущелья правого притока Терека - р. Армхи. Проведенные нами специальные исследования показали, что она широко распространена здесь на северных и отчасти южных склонах Бокового хребта в междуречье Терека и Аргуна, приблизительно до линии, соединяющей окр. сел. Терского и Советского на севере (рис. 42, 2). В северо-восточной Турции и Аджарии ареал рассматриваемой формы почти повсеместно перекрывается с ареалом L. s. paruula, а на южных и северных склонах - с ареалом L. caucasica.
Географическая изменчивость. Исследовались выборки из 5 популяций, расположенных в различных частях ареала подвида (табл. 17). На южных склонах Большого хребта с запада на восток исследованные популяции разделены расстоянием 120 и 260 км, тогда как крайние южная и северная выборки (в Аджарии и Чечено-Ингушетии) расположены на расстоянии 270 км друг от друга. Отметим также, что популяция в Чечено-Ингушетии в настоящее время изолирована на северном склоне Главного хребта и оторвана от основной части ареала, лежащего на юге. Характер изменчивости исследованных выборок представлен на рис. 43. Отвлекаясь от аджарской и чечено-ингушской популяций, можно видеть, что на южном склоне Главного хребта подавляющее большинство признаков фолидоза обнаруживает четкую клинальную изменчивость в сторону возрастания с запада на восток, что подтверждается также не включенным в таблицу дополнительным анализом промежуточной выборки из окр. Пасанаури в ущелье Белой Арагвы.
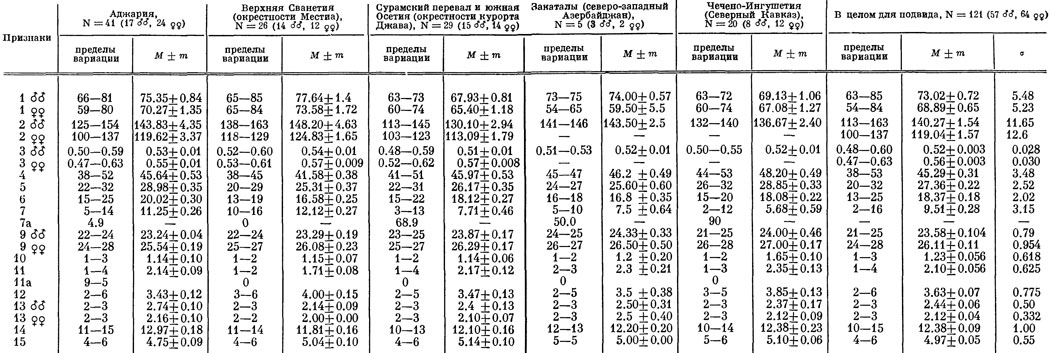
Таблица 17. Географическая изменчивость Lacerta rudis rudis
С другой стороны, отдельные признаки обнаруживают также клинальное изменение с юга на северо-восток (от Аджарии через Сурамский хребет в Чечено-Ингушетию).
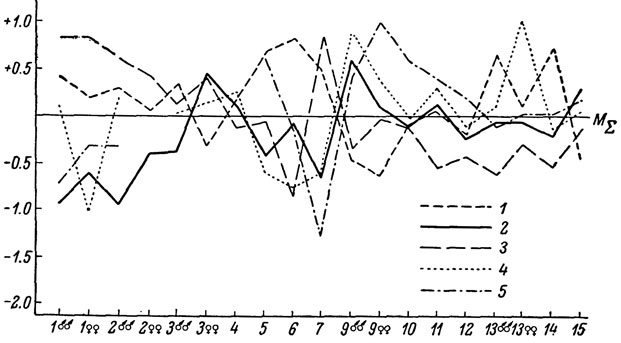
Рис. 43. Сводный профиль изменчивости L. r. rudis: 1 - Аджария; 2 - Сурамский перевал; 3 - Верхняя Сванетия; 4 - Закатали; 5 - Чечено-Ингушетия
Как и у многих других форм скальных ящериц, максимальные размеры тела (признаки 1 и 2) наблюдаются у ящериц, обитающих на большей высоте над уровнем моря. Следует также отметить, что ящерицы из Верхней Сванетии, помимо крупной величины, отличаются при жизни довольно яркой голубой окраской горла, особенно интенсивной у взрослых самцов.
Дерюгин (1901) и Боденгеймер (Bodenheimer, 1944) уже отметили чрезвычайно высокую изменчивость ряда признаков у изученных ими ящериц из окр. Трабзона, что подтверждается также и на исследованном нами материале из ряда пунктов между Трабзоном и Синопом. Анализ этого материала показывает, что на Черноморском побережье северо-восточной Турции существует хорошо выраженная зона гибридизации между L. rudis и малоазиатской формой L. s. lantzicyreni, ареалы которых отчасти перекрываются на южных предгорьях Восточно-Понтийского хребта.
Нужно также отметить, что экземпляры с Черноморского побережья Турции по ряду показателей фолидоза заметно отклоняются от особей из Аджарии и с Главного хребта, что позволяет допускать существование здесь особого подвида, отличного от номинальной формы. Об этом свидетельствуют и данные Дерюгина (1899), что скальные ящерицы из окр. Трабзона отличаются чрезвычайно яркой желтой окраской нижней стороны тела, чего никогда не наблюдается у L. rudis из Закавказья.
Сравнительные замечания. Бедряга (Bedriaga, 1886) описал свою Lacerta depressa var. rudis из окр. Батуми, а в дальнейшем Мегели (Mehely, 1909) и Никольский (1913, 1915) распространили это
название также на ящериц из Аджарии, северо-восточной Турции и южных склонов Главного Кавказского хребта. Несмотря на то что типовые экземпляры Podarcis depressa Camerano, 1878 (или по крайней мере некоторые из них) происходят из окр. Трабзона, вопрос о существовании на Черноморском побережье Турции L. s. rudis длительное время оставался спорным. Хотя Лантц и Цирен (Lantz et Cyren, 1936) выразили сомнение в принадлежности к этой форме нескольких просмотренных ими экземпляров из Трабзона, Боденхеймер (Bodenheimer, 1944) на составленной им карте указывает ее для целого ряда пунктов на побережье между Синопом, Трабзоном и Ризе. Для окр. Трабзона эта ящерица в последнее время была указана и нами (Даревский, 19656). Четкие морфологические отличия L. rudis, также как отсутствие промежуточных форм при совместном ее обитании с L. caucasica, L. mixta, L. armeniaca, L. s. brauneri, L. s. daghestanica и L. s. parvula, убедительно свидетельствуют в пользу видовой самостоятельности рассматриваемой формы.
Исследованный материал. Грузия: ЗИН, № 3382(1), Черная Арагва; № 13226 (1), Лагодехи; № 17745 (16), Сурамский перевал; № 17796 (3), Горная Тушегия, Омало; № 17814 (23), ущелье Ингури у впадения р. Накры; № 17875 (10), Местиа, Верхняя Сванетия; ГМГ (7), Пасанаури; (4), Леберде, Гегечкорский р-н; (6), Нижняя Сванетия, Лентехи; (2), Джвари, Цаленджихский р-н. Аджария: ЗИН, № 13500 (3), Зеленый Мыс близ Батуми; № 13501 (4), Зеленый Мыс; № 17515 (9), Шуахевский р-н; № 17517 (5), Чвана, Шуахевский р-н; № 17518 (13), Кеда, дорога на Мериси; № 17522 (5), р. Чаквис-Цкали, Кобулетский р-н; № 17523 (8), Кеда, дорога на Октомбери. Южная Осетия: ЗИН, № 17733, окр. курорта Джава. Азербайджан: ЗИН, № 17838 (4), Закатальский заповедник, р. Котехчай. Чечено-Ингушетия: ЗИН, № 15607 (2), Солш, Горная Ингушетия; № 17878 (13), ущелье р. Армхи, 8 км выше впадения в Терек; № 17882 (4), ущелье р. Аргуна, 4 км выше сел. Советского. Турция (северо-восточная): ЗИН, № 10688 (6), р. Ичхан-Су, Артвинский вил.; № 10689 (5), Кварцханы, Артвинский вил.; № 10690 (2), Ипхреул-Су, Артвинский вил.; № 10693 (5), р. Ичхан-Су, Артвинский вил.; № 10818 (1), Салолеттопасский пост. Артвинский вил.; № 13824 (1), Дзансуль, Артвинский вил.; № 13446 (1), Дзансуль, Артвинский вил.; № 17971 (1), Трабзон; ЕИМВ, № 18385, 2, 4 (2), Мериемана, близ Трабзона; № 18384 (1), хр. Ялнызчам, восточный склон; ЗМФ, № 11981 (1), Трабзон; № 12004 (1), Синоп; № 12008 (1), Синоп; № 12009 (1), Синоп.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://herpeton.ru/ 'Герпетология - о пресмыкающихся и земноводных'